Bilde 1: Egyptisk fruktfladder, av Mickey Samuni-Blank (eget arbeid) [ CC BY-SA 3.0 ], via Wikimedia Commons.
Vi har sett at medforfatter av en ny evolusjonsbok: Dennis Venema i 'Adam og Genomet', er påvirket av den mytiske krigføringsmetaforen (her og her) og gjør feilaktige argumenter om at det fossile beviset støtter evolusjosteorien (her). Når det gjelder 'krigførings'-metaforen, forkynner boken den falske historien om at det grunnleggende spørsmålet om Galileo-affære fra det 17. århundre var "den nye vitenskapens sannhet og dens oppfattede trussel mot bibelsk autoritet." Som vi så, er dette den falske evolusjonære overføring av historien. Krigførings-metaforen er en myte, og Galileo affæren er kanskje favoritteksemplet for {mange-oversetters tilføyelse} evolusjonister.
Når det gjelder det fossile beviset -her, som viser at arter opptrer brått i lagene), benytter boken to feilaktige argumenter: at evolusjon er nødvendig for at vitenskapen skal virke i det hele tatt (det filosofiske argumentet "intellektuelle nødvendighet") og bruken av tilfeldig design som alternativet til evolusjon (et teologisk argument).
Nå går vi videre til et annet emne: ekkolokalisering. Dette var av særlig interesse for Hunter siden han har brukt ekkolokalisering som et eksempel på hvordan evolusjon feiler. Da han så at Venema appellerte til ekkolokalisering for å argumentere for evolusjon, var han opptatt av å se hva han måtte si. C. Hunter leter alltid etter gode argumenter for evolusjon, men fant ingen her. Nedenfor oppsummerer han de fem forskjellige grunnene til at ekkolokasjon ødelegger for evolusjonsteorien. Til slutt vender han seg til Venema's argument, om det kan kalles det. Det vi vil se er at hans argument mislykkes totalt. Venema klarer ikke å løse noen av problemene med ekkolokalisering, og han klarer ikke å presentere noen form for et positivt tilfelle som kan brukes til å overvinne de mange problemene.
Kompleksitet: Den opprinnelige sonarteknologien
De fleste er kjent med begrepet radar og sonar. Enkelt sagt, et reflektert signal brukes til å spore et mål. Men det folk flest er mindre kjent med, er de mange detaljer og komplikasjoner som et radar- eller sonarsystem må regne med. For eksempel må den overførte puls være veldig sterk fordi den vil svekkes med kvadratet av avstanden den reiser, og bare en liten brøkdel av den vil bli reflektert. Til slutt er retursignalet veldig svakt, så mens mottakeren er utsatt for det meget kraftige overførte signalet, må det da oppdage et retur signal mange størrelsesordener svakere. Tenk på å rope så høyt du kan, og lytt etter ekko av en mygg.
Dette er bare begynnelsen på de mange sonardesign problemene. Pulsfrekvensen, varigheten, intensiteten, tonehøyde er alle designparametere som påvirker hvor lite mål som kan oppdages, hvor langt det kan oppdages, hvor nøyaktig det kan spores og løses, og så videre. En avansert sonardesign kan variere disse parametrene for å optimalisere sporing.

Sonar-design må også vurdere hvordan man kompenserer for målbevegelse og den resulterende Doppler-effekten, feil refleksjoner fra virvar i miljøet, og hvordan man kan lede mot et bevegelig mål. Det er også mulighet for avbildning for å bestemme hvilken type mål det er.
Ikke overraskende, er det mange forskjellige sonar-design strategier. Avhengig av 'støy' i miljøet, typiske måltyper og så videre, kan ulike designstrategier fungere bedre.
Alt dette er det vi finner i naturens ekkolokasjonsdesign. Hvaler og flaggermus har utrolig effektive og nøyaktige sporingsegenskaper. Vi har utviklet sonar, men naturen hadde det hele tiden - den opprinnelige sonarteknologien. Faktisk er naturens design bedre enn vårt militærutstyr. Det er en grunn til at de blir studert så nøye.
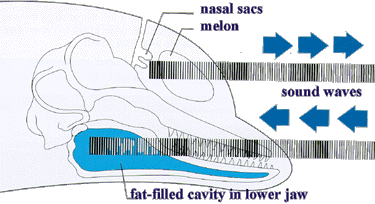
Bilde 2. Skisse av ekkkolokalisering (eksternt)
Kompleksitet på molekylær nivå
Vi har sett hvor komplisert ekkolokalisering kan være. Ikke overraskende er molekylære maskiner som bidrar til å få det til å skje, også svært komplekse. Prestin, et protein som er viktig for pattedyrs hørsel, er et transmembran-protein i de ytre hårcellene i sneglehuset (cochlea). Den fungerer som en frekvens-selektiv forsterker i et lydsystem som virker noe slikt som dette:
Når lyden kommer inn i øret, avbøyer den det ytre håret og forårsaker små mengder strekk eller kompresjon i ytre hårceller. Det er kanalproteiner som sitter i membranen til disse cellene, som er følsomme for en slik mekanisk belastning. Disse proteinene gir en tunnel (eller kanal) over membranen slik at ioner lett kan krysse, og den mekaniske belastningen kan føre til at kanalene åpnes.
Disse kanalene er nettopp laget for å tillate kun bestemte typer ioner å krysse. For eksempel tillater noen kanaler det positivt ladede kalium-ion å krysse, men ikke positivt ladede natrium-ioner og vice versa.
Når en kanal åpner, har ioner vanligvis en tendens til å krysse gjennom membranen (enten inn i cellen eller ut av cellen) fordi ione-koncentrasjonen ikke er jevn, og fordi det er spenning over membranen. Slike forskjeller i konsentrasjoner over membranen, og spenningen, holdes aktivt vedlikeholdt av cellen. De tjener som et slags batteri hvis energi kan tappes til enhver tid ved å åpne membrankanaler.
Når den innkommende lyden får visse kanaler til å åpne, forårsaker ionene som krysser en forandring i membranspenningen. I de ytre hårcellene oppfordrer denne spenningsendringen negativt ladede klor-ioner til å gå ut av cellen. De samhandler med prestin-proteinet, i membranen, for å forårsake en mekanisk deformasjon som resulterer i forlengelse av cellen.
 Med andre ord, den innkommende lyden som forårsaket at håret beveger seg, fører til enda større hårbevegelse, og dette tjener nettopp for å forsterke den innkommende lyden. Denne forsterkningen er større ved lave lydnivåer, slik den burde være.
Med andre ord, den innkommende lyden som forårsaket at håret beveger seg, fører til enda større hårbevegelse, og dette tjener nettopp for å forsterke den innkommende lyden. Denne forsterkningen er større ved lave lydnivåer, slik den burde være.
En av de interessante trekk ved dette systemet er hastigheten det opererer i. Tydeligvis, for å forsterke lyd må du reagere så fort som endringene i lyd forekommer. Proteinmotorer bruker ofte kjemisk energi (som splittelsen av ATP-molekylet), men det ville være for sakte for øret sitt lydsystem. I stedet bruker prestin membranens spenning. Denne elektriske energien kan brukes mye raskere og prestin opererer i mikrosekunds hastigheter. Her er hvordan en artikkel oppsummerte systemet:
"Den utsøkte høysensitivitet og frekvensselektivitet ved pattedyrs høreorgan stammer fra en mekanisk amplifikasjonsmekanisme som ligger i Corti-organet, følelsesorganet for hørsel i pattedyr. Gevinsten fra denne forsterkningen kan nå så høyt som tusenfold; Den er høyest ved lave lydnivåer og reduseres gradvis med økende lydenergi."
Evolusjonen har ingen forklaring på opprinnelsen til dette systemet utover ubegrunnet spekulasjon, og dette er bare starten på de mange molekylære maskiner bak ekkolokasjonssystemene som finnes i naturen.
Ekkolokaliserings-design ikke kongruent med evolusjons treet.
Bilde 3. Skisse av ekkkolokalisering (internt)
Det fremgår ikke at tilfeldige mutasjoner er årsaken til systemer som ekkolokalisering i flaggermus og hvaler. Selv om dette er et enormt problem for evolusjonsteorien, er det ikke den eneste. Som diskutert ovenfor er det mange forskjellige typer ekkolokasjonsdesign. Evolusjonen vil forutse at arter som antas å være nær naboer på det evolusjonære treet, vil dele lignende ekkolokasjonsdesign. Med andre ord bør ekkolokaliserings-design være kongruent med det evolusjonære treet. Men det er de ikke.
Mens Darwin hevdet at det evolusjonære treet forklarte naturens design i stedet for tilholdssted, følger naturens ekkolokaliserings-design den nøyaktig motsatte regelen. Her er hvordan en artikkel beskriver det:
"Dyrets habitat er ofte viktigere for å forme sin oppbygning av kallesignal (call design) enn deres evolusjonære historie.
Dette er en enorm falsifisering av en nøkkelforutsigelse i evolusjonsteorien.
Konvergens på det morfologiske nivået
En konsekvens av denne falisifiseringen er at evolusjonistene må konstruere svært kompliserte fortellinger for opprinnelsen til ekkolokalisering. For eksempel, hvis evolusjonsteorien er sann, må vi tro at den utrolige ekkolokasjonsevnen som finnes i enkelte flaggermus, oppsto flere ganger, ved å utvikle seg selvstendig. Det er ikke lett for evolusjonister å forklare. Hvordan kan slike forbløffende designdetaljer gjenta seg via blind biologisk variasjon? (nei, naturlig utvalg hjelper ikke)
Men dette konvergensproblemet går langt utover flaggermusene. Hvaler og flaggermus deler noen forbløffende likheter i hvordan de sporer sine byttedyr. Men hvis evolusjonsteorien er sann, må vi tro at deres vanlige forfedre ikke hadde noen av disse evnene. Så i helt forskjellige deler av verden, i helt forskjellige miljøer, må tilfeldige mutasjoner i disse forskjellige artene selvstendig ha konstruert de samme ultrakomplekse designene. Som en rapport forklarte:
"Selv om de utviklet seg separat over millioner av år i forskjellige verdener av mørke, bruker flaggermus og tannhvaler overraskende lik akustisk oppførsel for å lokalisere, spore og fange bytte ved hjelp av ekkolokering, den biologiske ekvivalenten til sonar. Nå har et team av danske forskere vist at den akustiske oppførelsen til disse to dyrene under jakt er nifst like."
 Hvis evolusjonsteorien er sann, ville flaggermus og hval ha utviklet seg selvstendig i millioner av år. Og likevel konstruerte de begge en sonar-evne som innebærer overføring av høye signaler mens de mottar utrolig svake signaler, justering av signal-parametrene i sanntid, behandling av mottatte signaler og så videre. De deler til og med samme utvalg av ultralydsfrekvenser:
Hvis evolusjonsteorien er sann, ville flaggermus og hval ha utviklet seg selvstendig i millioner av år. Og likevel konstruerte de begge en sonar-evne som innebærer overføring av høye signaler mens de mottar utrolig svake signaler, justering av signal-parametrene i sanntid, behandling av mottatte signaler og så videre. De deler til og med samme utvalg av ultralydsfrekvenser:
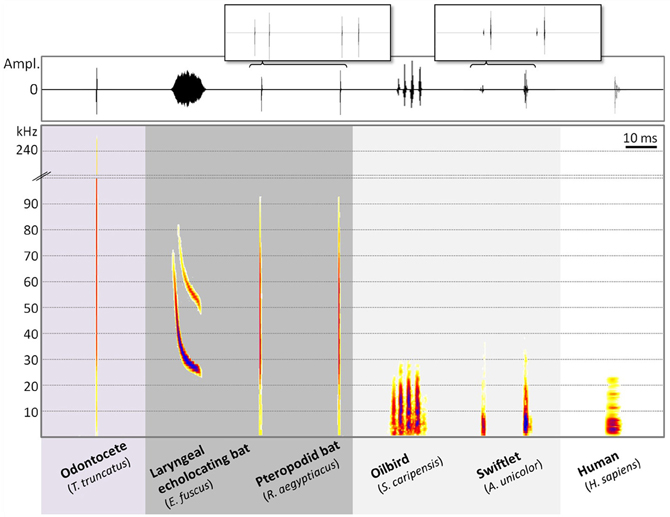
"Flaggermus og tannhvaler (som inkluderer delfiner og niser) hadde mange muligheter til å utvikle ekkolokeringsteknikker som avviker fra hverandre , siden deres nærmeste stamfar ikke var i stand til ekkolokalisering. Likevel, som forskere har visst i årevis, er flaggermus og tannhvaler avhengig av samme rekkevidde av ultralydfrekvenser, mellom 15 og 200 kilohertz, for å jakte på byttet.
Bilde 4. Ekkolisering -amplitude
Og den likheten er til tross for de forskjellige miljøene:
"Denne overlappingen i frekvenser er overraskende fordi lyden beveger seg omtrent fem ganger raskere i vann enn i luft, noe som gir tannhvalene betydelig mer tid enn flaggermusene for å velge å snappe opp et potensielt måltid."
Men det er ikke alt. Flaggermusen og hvalen bruker også lignende strategier for å justere signalene sine mens de går for ett byttedyr:
"Flaggermus øke antall samtaler per sekund (som forskere kaller en "summe rate") mens de er på jakt etter byttedyr. Hvaler ble antatt å opprettholde en jevn hastighet av samtaler eller klikk, uansett hvor langt de var fra et mål. Men den nye undersøkelsen viser at villhvaler også øker antallet av samtaler eller klikk under en drap - og at hvalens 'summe-frekvens' er nesten identiske med flaggermusens , ved omtrent 500 'klikk' per sekund."
Det er et annet eksempel på en kompleks designutvikling som en bare kan spekulere om, og igjen feiler det evolusjonære treet i å forutsi dets mønster.
Konvergens på molekylær nivå
Ikke bare er utrolig ekkolokaliserings-konvergens tydelig på det morfologiske nivået, det ses også på molekylært nivå. For eksempel er prestin-proteinene i visse flaggermus og hvalarter mer like enn evolusjonsteorien forventer. Det massive prestinproteinet har for mange aminosyrer som samsvarer mellom disse artene. Hvis man skulle bygge et evolusjonært tre på grunnlag av prestin-sammenligninger alene, så ville flaggermusen og hvalen bli gruppert sammen, og det kan ikke være riktig.
Dette faktum alene trenger ikke være et problem for evolusjonister. De sier bare at prestin er under påvirkning av sterk seleksjon. Med andre ord er det sterke funksjonelle begrensninger på prestin som krever mer likhet, selv mellom fjerne arter, enn vi vanligvis finner i proteiner.
Bilde 5. Prestin-proteinet
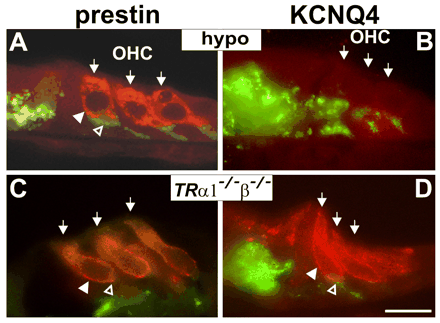
Spesielt identifiserte forskere ni aminosyrer i prestin som synes å være ansvarlige for den altfor konsekvente hval-flaggermus 'matchen'. De ni aminosyrene må være under svært sterkt utvalg. Hvis én av dem muterte da ville bio-sonarsystemet ikke fungere bra. Flaggermusen eller hvalen ville ikke overleve, og det er derfor vi ikke observerer slike endringer. Det er slik naturlig valg fungerer.
Men hvis alle ni aminosyrene er påkrevd, hvordan snublet evolusjonen borti dette designet i utgangspunktet? Det ville være svært lite sannsynlig at de rette ni aminosyrene skulle oppstå via blinde mutasjoner, samtidig.
Men konvergensen til molekylære maskiner bak ekkolokalisering går langt utover prestin. Som en artikkel forklarer, er konvergens ikke en sjelden prosess begrenset til noen steder, men er i stedet utbredt. "
Som en evolusjonist innrømte -her , "Disse resultatene innebærer at konvergerende molekylær evolusjon er mye mer utbredt enn tidligere anerkjent." Og en annen innrømmet her at resultatene er forbløffende:
"Vi hadde forventet å finne identiske endringer i kanskje et dusin gener eller så, men å se nesten 200 er utrolig. Vi vet at naturlig utvelgelse er en kraftig driver for gensekvens-utvikling, men det er forbløffende å identifisere så mange eksempler hvor det produserer nesten identiske resultater i de genetiske sekvensene av helt urelaterte dyr.
Forbløffende. Mer om 'protein-evolusjon -nylonase'.
Venema: Hvorfor ekkolokalisering ikke er et problem
Dette bringer oss til Venemas argument for hvorfor ekkolokalisering ikke er et problem. Gitt de enorme vanskelighetene som gener gjennomgått - kortfattet ovenfor: hvordan eksakt finner Venema ekkolokalisering evolusjonsvennlig? Vi har sett på problemet med kompleksitet av ekkolokering, blant annet på molekylært nivå, problemet at ekkolokasjons-design er uoverensstemmende med det evolusjonære treet, og som et eksempel problemet med konvergens på både de morfologiske og molekylære nivåene. Sikkert nok ville ingen objektiv forsker finne bevis for evolusjon i naturens ekkolokasjons-design.
Bilde 6. Flaggermus-sonar
Ville de? Tro det eller ei, her er hva Venema skriver: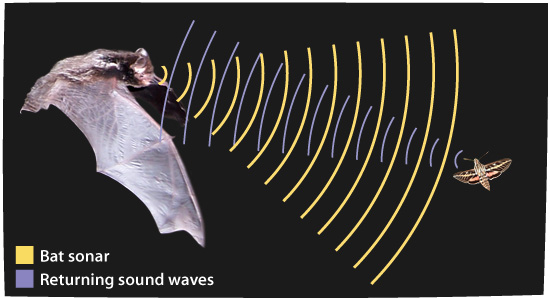
"Hvis du noen gang har snublet gjennom et bekmørkt rom og trakk deg like før du kolliderte med en vegg eller et annet objekt, har du anvendt din (veldig rudimentære) sans for ekkolokalisering. Det du oppdaget (selv om du kanskje ikke selv har bevisst oppfattet det) var at lydbølgene reflekterte fra objektet i veien. Alle pattedyr kan gjøre dette , men de fleste (lik oss) gjør det veldig dårlig. Vi må være svært nær det aktuelle objektet før det endog er mulig for oss å legge merke til reflektert lyd, og like sannsynlig, vil vi ikke gjøre det, og vi vil krasje tåa eller verre.
Som det viser seg, er sjøpattedyr-ekkolokalisering en spesifikt fininnstilt hørselsfornemmelse som er basert på de samme gener som brukes til å høre hos andre pattedyr. Et nøkkel-gen som brukes til å høre hos alle pattedyr kalles "prestin" -genet, et protein som er involvert i spesialiserte strukturer i pattedyrøret som vibrerer i respons på lydbølger. I hvaler er prestin-genet innstilt på ultralyd-frekvensene som er bedre egnet til ekkolokalisering. Denne tuningen krevde bare noen få aminosyre-endringer i proteinet - en mengde forandring lett innenfor rekkevidden av den slags molekylære endring vi så for insulingenet i forskjellige pattedyr. Denne endring i prestin-genet for å tune det til ekkolokalisering var så lett å oppnå, det virket som om nesten identiske endringer skjedde uavhengig i linjen som førte til moderne flaggermus, som også bruker en prestin innstilt på ultralydfrekvenser for ekkolokalisering. Så selv ekkolokalisering er ikke "ny" - den er også omformet fra en standard pattedyr-sans for hørsel."
Dette er en komplett katastrofe. Venema's sammenligning av ekkolokalisering med sin forestillede evne til å unngå en vegg i et mørkt rom, hans endrende konvergens til en dyd, hans fremlegging av ekkolokalisering som "lett å oppnå", resultatet av bare "flikking" og ikke noe nytt, men ganske enkelt en remodellering av "standard pattedyrs følelse av hørsel" er alle standard evolusjonær 'krumkastings'-logikk.
 Dette er den evolusjonerende "bare føy til vann" -synet på biologi hvor du legger til et par mutasjoner og 'poff', så har du ekkolokalisering. Men som vi så over, er ekkolokalisering ikke i det hele tatt sammenlignbart med "standard pattedyr" hørsel. Den passer ikke til det evolusjonære treet, og konvergensen er forbløffende og helt uventet og uforklarlig.
Dette er den evolusjonerende "bare føy til vann" -synet på biologi hvor du legger til et par mutasjoner og 'poff', så har du ekkolokalisering. Men som vi så over, er ekkolokalisering ikke i det hele tatt sammenlignbart med "standard pattedyr" hørsel. Den passer ikke til det evolusjonære treet, og konvergensen er forbløffende og helt uventet og uforklarlig.
Venemas forsøk på å bortforklare ekkolokalisering som et standardresultat for evolusjon er 'ikke engang feil' -{den er på feil jorde -oversetters kommentar.}
C. Hunter har fulgt ekkolokaliseringsforskningen i mange år. Han skriver om det, og inkluderer ofte det i presentasjoner. Han diskuterer de ulike måtene som ekkolokasjonsbeviset motsetter seg evolusjonsteorien. Så hvorfor skulle det være en seksjon om dette emnet i denne boken som fremmer evolusjon? Han lurer på: 'Har jeg savnet noe? Er det noe grunnleggende aspekt ved ekkolokalisering han har gått glipp av? Er det en ny artikkel jeg har oversett, som snur opp ned på det store forskningstmaterialet?' {Oversetters forslag er at innslaget var ment som et forsøk på å forsvare evolusjonsteorien på dette punktet, -uten å lykkes.}
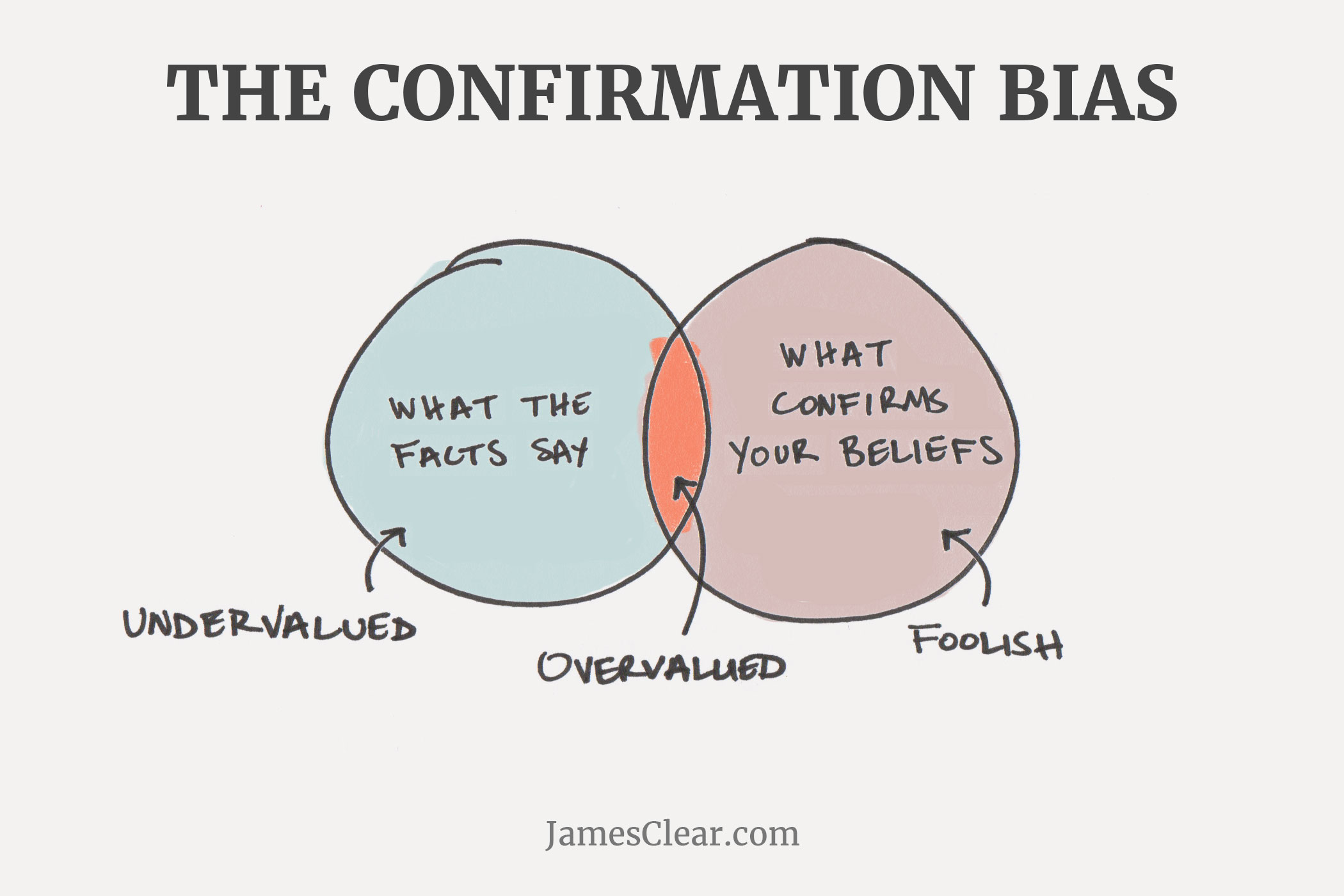
Bilde 7. Eks. på bekreftelses-avvik
Nei, da forfatteren (C. Hunter) leste innlegget, skjønte han raskt at det ikke var noe mer enn den vanlige evolusjonære 'det-bare-er-slik-historien'. En helhetlig ignorering av veletablert vitenskap, en omfavnelse av forestilte tankeeksperimenter som ikke gir mening, og en ytterst uvitenskapelig konklusjon.
Oversatt av Asbjørn E. Lund
(Bildene sto ikke i opprinnelig artikkel, evt. untatt det 1., og er satt inn av undertegnede, se lenke i Bilde-nr)
 Ekkolokalisering -feilaktige påstander
Ekkolokalisering -feilaktige påstander