Livet er godt
(Utvalgte deler fra Kap. 6 i 'Undeniable' av Douglas Axe, Harper One)
Etter å ha etablert at vi alle kan bedrive allmenn vitenskap, og at vi ikke blindt skal akseptere det profesjonelle vitenskapsfolk tenker om livet, er neste steg å finne ut hva vi tenker om livet. Livet kan noen ganger uttrykkes ved en wow-faktor, men det som gjør livet unikt attraktivt for oss synes dypere enn denne 'wow-faktoren'. Jeg tror det er noe nærmere hensikt. F.eks. tornadoer kan score høyt på 'wow-faktor', p.g.a. deres enorme potensiale. Mens tornadoer gjør 'sin greie' med stor intensitet, så forsøker de ikke gjøre hva de utfører, fordi tornadoer har ikke noe sinn eller bevissthet. Edderkopper forsøker på sin side å fange insekter, selv om disse prøver å komme løs fra sin til-fangetagers nett.
Hvorvidt aktivitet fra mye enklere livsformer, som f.eks. en amøbe, involverer bevissthet, beror mye på gjettverk. Forfatteren mistenker en amøbe er mer å sammenlikne med en maskin, enn en puma i så måte. Om han skulle velge en maskin for å illustrere en amøbe, ville det være såkalt krypende vaskeroboter, som benyttes til å renske svømmebasseng. De leter stadig etter rester/skrap i stedet for mat, men bevegelsene er nesten livaktige i sin kompleksitet. Basseng-roboter skulle overbevise oss om at at en ting ikke trenger å være bevisst, for at vi skal fornemme hensikt når vi observerer den. De som observerer dem, vil etter noe tid komme til at om en summerer alle deres små bevegelser opp på et høyere nivå, så vil vi ane at hensikten med roboten er at den renser bassenget.
Det samme gjelder i enda høyere grad for livet. Et barn som ser en edderkopp lage et nett for første gang, og ser en hel masse bevegelser som ender i et helhetlig design. Det både synes og er virkelig 'gripende'. De små bevegelsene får mening, fordi de produserer et betydelig resultat, og vi kan ikke unngå overbevisningen at dette var et planlagt resultat. Det var en grunn til edderkoppens travelhet. Det er ikke alltid aktivitet legger igjen det inntrykket. Litt regn legger igjen mange små regndammer. Selv om det fortsetter til veier flommer over, så sitter vi ikke igjen med at det var hensikten. Regn bare forekommer, men livet forekommer ikke bare. Liv er så ulikt regn at vi kan ha nytte av et annerledes vokabular, for å snakke tydelig om det.
Travle helheter og hele prosjekter
I følge 'Oxford dictionary' er helhet 'en ting som er fullstendig i seg selv.' Edderkopper og basseng-roboter er hele i denne betydning, mens sanddynger, regndammer og tornadoer ikke er det. Handlinger som deler en sandhaug, gir oss sammenlignbare deler med originalen, bare mindre. Disseksjon av en edderkopp eller en basseng-robot etterlater oss deler, som ikke i det hele er sammenlignbare med helheten de kom fra. Det samme kan sies om et karbonatom eller om solen, begge har karakteristika som kommer bare ved å summere delenes. Men disse manifesterer ikke hensikt, på samme måte som en edderkopp eller basseng-robot gjør. Disse er eks. på en spesiell form for helhet, som manifesterer hensikt ved å overta og fullføre ett prosjekt. Det innebærer ikke at ting utenfor denne klassen av helhet, ikke er tiltenkt eller ordinære. Poenget er at ingen av de ekskluderte tingene arbeider, slik en edderkopp eller bassengrobot gjør, som om de har en hensikt knyttet til seg. For å kjennetegne den nevnte klassen av helhet, skal vi betegne den som 'travle helheter'. En travel helhet er da, en aktiv ting som får oss til å oppfatte hensikt, fordi den gir et resultat ved å bringe mange små ting eller omstendigheter sammen, på nøyaktig riktig måte. Resultatet er også et hele, som vi kan kalle et 'helt prosjekt', Så travle helheter er helheter som vi vil kalle hele prosjekter. Når vi ser fullførte 'hele prosjekter' ser vi hensikt , uansett hvordan det ble utført.
Når vi gjenkjenner en situasjon eller et objekt, som et helt fullført prosjekt, innser vi at fullførelsen krevde arbeid. Mer spesifikt ser vi at fagkyndig arbeid var påkrevd, arbeid som brakte riktige ting sammen på en rett måte. I vår erfaring krever evne, skjønnsomhet, evne til skjelne de rette fra de gale tingene. Slik skjelning krever i sin tur igjen kunnskap. Om vi erkjenner det, så slutter vi at en eller flere kyndige må ha stått bak arbeidet. Det følger naturlig fra design-intuisjonen vi har henvist til. Legg merke til at resultatet flytter seg fra resultatet, det fullførte prosjektet, til den som utførte det. Kunnskap og hensikt blir trukket inn slik at det ikke kreves at vi vet hvem som visste eller hadde noe til hensikt. Om vi ser en arbeidsrobot i virksomhet, vet vi at å takle slike prosjekter krever kunnskap, uten at roboten i seg selv har noen bevissthet om dette. I stedet erkjenner vi at en basseng-robot er et suksessfylt resultat av et mye mer imponerende helt prosjekt: designing og fremstilling av en fungerende basseng-robot. Så kunnskapen og hensikten vi fatter, når vi observerer basseng-roboten, er i siste omgang sporbar til menneskelig kunnskap og hensikt, uten at vi dermed vet hvordan den foregikk.
Travle helheter takler sine prosjekter ved å bryte dem ned i mindre prosjekter, på en organisert måte. Store prosjekt deles i mindre under-prosjekter. Så lenge underprosjektene er komplekse nok til å kvalifisere som hele prosjekter, sanser vi at deres fullføring drives ut av hensikt. Disse forestillingene er mer kjente enn de kan synes. F.eks. å vinne en tenniskamp kan være et helt prosjekt for en tennisspiller. Suksessen er kritisk avhengig av mange under-prosjekt. Én er tilførsel av oksygen og transport av CO2 fra blodet, som et helt under-prosjekt. Den travle helheten som har det overordnede ansvaret, er ett menneske, mens de travle prosessene som styrer fysiologiske prosesser, støtter hoved-prosjektet. Sammen med lungene i puste-prosjektet, er andre travle helheter som nervesystemet og mellomgolvet. Og som en kan forvente, prosjektene som blir fullført av hver av disse anatomiske travle helhetene, kan bli videre delt opp i underprosjekt tilordnet vev, og celler, så til subcelle og intercellulær samhandling, og endelig ned til livets molekyler.
Nå blir spørsmålet som interesser oss, hvorvidt noen hadde til hensikt at våre lunger og indre celler skulle takle sine respektive prosjekter, slik tennisspilleren har til hensikt å vinne matchen. Vi tenker ubevisst at celler og organer naturlig var ment å være slik, og en allmenn-begrunnelse kan tilbys for denne tendensen. Vi skal nå prøve å bygge en sak for å vise at en slik slutning til hensikt er korrekt. Vi skal prøve å se at det er mer å vinne ved å grunngi et mer tilfredsstillende syn på livet, enn ved å vise at Darwin tok feil, selv om begge deler tilsiktes.
Om laks og spekkhoggere
Forfatteren har kommet til at alt om laks, dreier seg om laks, og alt om spekkhoggere dreier seg om spekkhoggere. Som molekylærbiolog gjennom flere tiår, vet han likhetene mellom disse to er reelle og signifikante. Men når han ser dem i virksomhet, forsvinner denne teoretiske kunnskapen. F.eks. når han ser salthavslaks, kjempe seg opp gjennom elva de kom fra, for å nå fram dit de selv ble født. Deres oppdrag fortærer dem bokstavelig talt. De forsaker all mat, de ofrer kroppen sin i det de kaster seg over steiner, og kjemper seg oppover, for å bli foreldre til avkom de ikke kommer til å se. I deres kompromissløse bestemthet, gjør disse strålende skapningene det helt klart, at de gjør akkurat det de var ment å gjøre, selv om mange av dem dør før de oppnår det. Det er noe heroisk over det de er, og noe strålende over intensiteten det leves ut med.
Livet som Darwin så det
Om det er noe som er drivende i Darwins syn på livet, så er det enkelheten i hans kjerne-ide. Under det virvar av idéer som evolusjonsbiologi er blitt, er det ett kjerneprinsipp: organismer med evne til å reprodusere seg, bærer potensiale til å produsere etterkommere som er bedre i så måte. Denne vedvarende forbedring av reprodusere, synes bare å ha å kreve: i) at de fortsetter å utføre sin reproduksjon ikke-perfekt -at det er små feil (mutasjoner) som tidvis blir introdusert og ii) at i det minste noen av disse feilene forbedrer reproduksjonen, om enn bare litt. Vi skal nå betrakte hvorvidt denne forklaringen, om vi aksepterer den, skal forme vårt bilde av livet. Tatt i betraktning hvor ufarlig denne formodningen synes, så kan dybden av dens implikasjoner komme som en overraskelse. Vi overlates til å se de mange livsformer på samme måte som vi ser geologiske formasjoner, som ting i stadig endring. Om fjell ser permanente ut for oss, fordi de beholder sine formasjoner i lange tider, så vet vi at de konstant påvirkes av naturkrefter omkring dem.
I følge dette synet, må det ha vært en enkelt stamfar for alle dyr, -hvis avkom ble drevet av naturlig seleksjon i mange ulike retninger, lik løv spredt utover vannet av overførings-strømmer. Moderne dyr kan da ikke være annet enn nåværende lokalisering av disse spredte løvene. Hver tilsvarer et bilde i en langfilm, dagens snapshot. Størrelsesorden til de kumulative endringer kan forbløffe oss, i det vi betrakter den svimlende variasjon av dyr som kom fra en enkelt stamfar. Formodentlig vil etterkommerne av dagens edderkopper, hvaler og laks bli radikalt ulike, gitt så mye tilgjengelig tid igjen.
Selv om forfatteren personlig misliker denne idéen om livets flyktighet, så måtte en komme overens med den, dersom en var overgitt til ideen om naturlig seleksjon som livets skaper. Forfatteren blir konstant opptatt av motsigelsen mellom det han ser når han åpner øyene, og evolusjonsteorien. For ham ser livet meget ulikt ut i forhold til geologi. Geologi forstås best ved å gruppere objektene i noen relativt få store kategorier. Mens biologi på sin side synes å behøve en annen oppdeling. Om en seriøst følger en forståelse av livets distinkte variasjoner, så tvinger det oss til å oppgi ideen om at de fundamentalt sett er samme greia. Verken edderkoppen, laksen eller hvalen synes å passe den idéen. Hver av dem er slående interessante og fullstendige, helt overgitt til å være det de er. Hver av dem vil fullføre livsforløpet heroisk ved død eller utryddelse, men ikke ved overgivelse til krefter som vil gjøre dem til noe annet.
Bilde 1. Eks. på livets opprinnelse
 Perfeksjon og dens kritikere
Perfeksjon og dens kritikere
Temaet med overgivelse tar helhets-forestillingen til ett nytt nivå: Den hinter om at noen helheter er det de er , fordi de burde være slik. Det er som de er representasjoner av noe sannere og mer betydelig enn noen midlertidig, fysisk representasjon. Idéen her er at noen organismer er så gode, at de ikke kan være særlig annerledes enn de er. Det innebærer ikke at de er så gode at de måtte eksistere. Omfanget av menneskelig kreativitet er prydet med flere eksempler, en perfekt musikalsk komposisjon, ett skjønt malerie, ett perfekt matematisk bevis, tidløse skatter for å betraktes, men ikke for å omarbeides. Livet er den vesentligste representasjon av denne idéen, uten rivaler blant menneskelige bedrifter. Gamle lærebøkers forsøk på å definere livet, som at det er en selvoppholdende, ikke-likevektstilstands prosess, basert på karbon-kjemi og drevet av tilstrømning av solenergi, kan fort glemmes. Den resonerte ikke hos noen som funderte over livet. Nei, livet må være noe mye rikere, umåtelig mye mer verdt vår oppmerksomhet. Livet er et mysterium og ett mesterstykke, en overflødig rikdom i perfekte komposisjoner. Du og jeg er blant dem, for å nyte det en kort stund.
Alle 'senser' livets dype under. Det synes for overveldende til å bli oversett. Like tydelig er spenningen mellom dette opphøyde syn på livet og forklaringen som tilbys av Darwin. Hans idé om at livet vandrer fra en variasjon til den neste, uten å slå seg til ro, overgitt til den blinde kraften i naturlig seleksjon, er inkompatibelt med forestillingen at fysiske former er uttrykk for noe dypere, noe ubevegelig, noe perfekt. Så hvordan reagerer en person som er motvillig til å oppgi det darwinistiske alternativet på dette opphøyde synet på livet? Det best kjente alternativet er vel å prøve å degradere det høye synet. En understreker aspekter ved livet som vi alle vet, ikke er gode eller rette. Axe sitt syn går imidlertid dypere enn det. Han fornekter ikke at nåværende status for livet, er problemfylt på mange områder. Hans poeng er imidlertid å bekrefte at noe iøynefallende godt, klart er mulig å skjelne, selv gjennom tilslørende problemer.
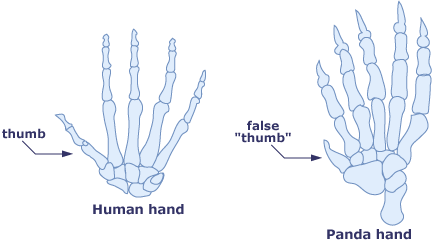 En annen måte å nedgradere livet på, er å anta at rollen til som 'bio-kritiker', en som ser etter feil i livets design. Som ett eksempel har den store pandaen en framstikkende ben i vristen, som har en tommel-lignende rolle, i det den hjelper den store bjørnen å få tak i bambus. Det er et faktum at dette lille uavhengige benet (radial sesamoid), ikke er en skikkelig sammenføyd tommel, lik vår. Dette har ført til at noen kritikere har påstått at ingen skikkelig designer ville 'gjort det slik'. Ikke overraskende argumenterer andre at det er godt design. Axe vurderer kritikerne mer enn pandaen. Ingen av kritikerne, selv hvor seriøst de mener det, har noe dypt grep om designprinsipper og utvikling som ligger under slike uavhengige ben, 'tomler' eller pandaer. Selv i topp-ressurssentre i verden finnes ingen 'skjelett-ingeniører', som vet hvordan en best kan sette bakbein på igler. Vår uvitenhet hittil om disse 'hvordan' spørsmålene, gjør oss også uskikket til seriøst å engasjere oss i 'hvorfor' spørsmålene.
En annen måte å nedgradere livet på, er å anta at rollen til som 'bio-kritiker', en som ser etter feil i livets design. Som ett eksempel har den store pandaen en framstikkende ben i vristen, som har en tommel-lignende rolle, i det den hjelper den store bjørnen å få tak i bambus. Det er et faktum at dette lille uavhengige benet (radial sesamoid), ikke er en skikkelig sammenføyd tommel, lik vår. Dette har ført til at noen kritikere har påstått at ingen skikkelig designer ville 'gjort det slik'. Ikke overraskende argumenterer andre at det er godt design. Axe vurderer kritikerne mer enn pandaen. Ingen av kritikerne, selv hvor seriøst de mener det, har noe dypt grep om designprinsipper og utvikling som ligger under slike uavhengige ben, 'tomler' eller pandaer. Selv i topp-ressurssentre i verden finnes ingen 'skjelett-ingeniører', som vet hvordan en best kan sette bakbein på igler. Vår uvitenhet hittil om disse 'hvordan' spørsmålene, gjør oss også uskikket til seriøst å engasjere oss i 'hvorfor' spørsmålene.
Bilde 2. Pandaens 'falske tommel' er benyttet som 'evolusjonsbevis'
Et bedre alternativ for folk som ikke er klar for å skilles med Darwins teori, er å omfavne livets storhet, i håp om at dette en gang skal vise seg forklarlig i darwinistiske termer. Dette alternativet har den betydelige fordelen av å bekrefte vårt høye syn på livet. Likevel har det utfordringer som kan sammenlignes med å få en kvadratisk påle til å passe i et rundt hull. Om naturlig seleksjon ikke bare er 'mester-formeren', men også en uopphørlig 'felespiller', som Darwin tenkte, så når aldri darwinismen en overbevisende avslutning (1). For Darwin ville tanken om alle de ulike evolusjonære linjer som avsluttes ved ender som er for gode til å endres, være like vanskelig å akseptere som tanken på uendrede eksterne forhold. Lokale økosystemer og klima utsettes for stadige endringer, som innebærer at forhold aldri slår seg endelig til ro. {At noen organismer implisitt har evne til å tilpasse seg endringer i omgivelsene, skjer mye raskere enn darwinistiske mekanismer kan forklare -oversetters tilføyelse.} Bekreftelsen av at det er noe unikt overbevisende ved levende organismer som vi ser dem nå, er en bekreftelse på ferdigstilling. Det fornekter idéen at livets design kan sammenlignes med løv som driver på en dam, eller alltid skiftende fjellformasjoner lik bilder i en film. Darwinistiske tilhengere synes å måtte velge mellom å tro på sin teori eller sine øyne.
 For å forstå dette dilemmaet tydeligere, kan vi forsøke å se for oss en troverdig evolusjonær forløper til alle moderne dyr. Etter å ha nedstammet fra de enkleste livsformer, ville denne skapningen bare hatt de mest grunnleggende basis-egenskaper som finnes i alle dyr. Den ville skille seg fra planter ved å mangle celle-vegger og fotosyntese. Blant moderne dyr, kan en tenke på svamper, som noe som kunne tilsvare en slik beskrivelse. Om denne fortidige svampen ville produserte f.eks. en moderne spekkhogger, gjennom en lang rekke av overgangsformer, kan vi spørre: Hva var det som drev denne forbløffende endring av dyreformer langs denne spesielle avstamningslinjen? Det synes bare å være to muligheter: Enten var det livsforholdene som bestemte utviklingsforløpet, og/eller naturlig seleksjon leverte det som dagens forhold favoriserte. Det første alternativet impliserer at fortidige svamper og nåværende spekkhoggere er forbundet med en suksesjon av tilpassede dyreformer, mens den andre impliserer en oppadgående bevegelse, fra de enkle fortidige til de overlegne moderne.
For å forstå dette dilemmaet tydeligere, kan vi forsøke å se for oss en troverdig evolusjonær forløper til alle moderne dyr. Etter å ha nedstammet fra de enkleste livsformer, ville denne skapningen bare hatt de mest grunnleggende basis-egenskaper som finnes i alle dyr. Den ville skille seg fra planter ved å mangle celle-vegger og fotosyntese. Blant moderne dyr, kan en tenke på svamper, som noe som kunne tilsvare en slik beskrivelse. Om denne fortidige svampen ville produserte f.eks. en moderne spekkhogger, gjennom en lang rekke av overgangsformer, kan vi spørre: Hva var det som drev denne forbløffende endring av dyreformer langs denne spesielle avstamningslinjen? Det synes bare å være to muligheter: Enten var det livsforholdene som bestemte utviklingsforløpet, og/eller naturlig seleksjon leverte det som dagens forhold favoriserte. Det første alternativet impliserer at fortidige svamper og nåværende spekkhoggere er forbundet med en suksesjon av tilpassede dyreformer, mens den andre impliserer en oppadgående bevegelse, fra de enkle fortidige til de overlegne moderne.
Bilde 3. Svamper -kan differensiere cellulært- til hva?
Begge disse scenariene har sider ved seg som er verdt å ta opp. Om forholdene er i førersete, sier vi at livet hovedsakelig er åpent, på grensen til det usammenhengende, for enten å være en svamp eller en spekkhogger. Eller hvilke som helst mindre endringer, for å dekke det ikke så lille gapet. Om vi derimot sier at naturlig seleksjon er dominant, så kommer vi ubehagelig nær å 'personlifisere evolusjon'. Som om den hadde visjon til å endre en enkel svamp, gjennom lange perioder av vanskelig 'oppvekstkår', da den 'visste hvor godt sluttresultatet ville bli'.
Hvorfor proteiner ikke utvikler seg (lengre)
Bilde 4. Protein-foldinger skjer 3-dimensjonalt
 Det store spørsmålet her, er imidlertid i hvilken grad livet er åpent for evolusjonær omdannelse. Svaret som har dukke opp med økende klarhet i senere år, ville ha overrasket Darwin. Noen nøkkel-faktorer tar oss tilbake til emnet med proteiner. For å forklare hvordan naturlige proteiner, med deres eksplisitte funksjoner kunne ha dukket opp ved uhell, er en monumental utfordring. Denne utfordringen kan deles i ett ekstremt aspekt, og ett mindre ekstremt, som begge er hovedhindringer for evolusjonsteorien. Den mer ekstreme utfordringen er å forklare hvordan mutasjoner og seleksjon kunne ha produsert komplett nye strukturelle tema for proteiner, som kalles foldinger. Det mindre ekstreme alternativet er å forklare hvordan mutasjoner og seleksjon kunne ha produsert funksjonelle variasjoner på eksisterende folder.
Det store spørsmålet her, er imidlertid i hvilken grad livet er åpent for evolusjonær omdannelse. Svaret som har dukke opp med økende klarhet i senere år, ville ha overrasket Darwin. Noen nøkkel-faktorer tar oss tilbake til emnet med proteiner. For å forklare hvordan naturlige proteiner, med deres eksplisitte funksjoner kunne ha dukket opp ved uhell, er en monumental utfordring. Denne utfordringen kan deles i ett ekstremt aspekt, og ett mindre ekstremt, som begge er hovedhindringer for evolusjonsteorien. Den mer ekstreme utfordringen er å forklare hvordan mutasjoner og seleksjon kunne ha produsert komplett nye strukturelle tema for proteiner, som kalles foldinger. Det mindre ekstreme alternativet er å forklare hvordan mutasjoner og seleksjon kunne ha produsert funksjonelle variasjoner på eksisterende folder.
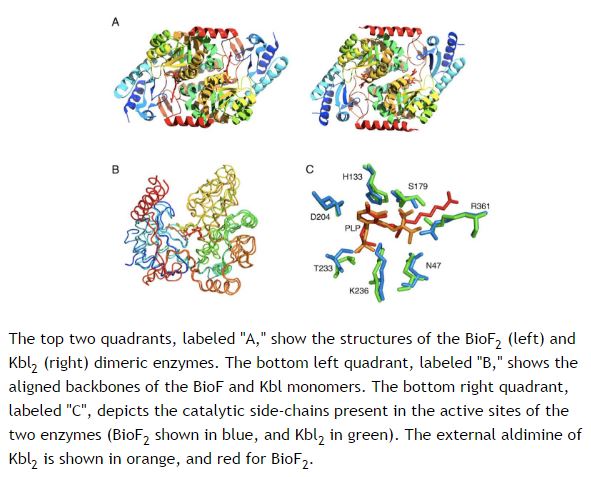
Dette prosjektet gikk Axe og hans kollega Ann Gauger inn på. De valgte å arbeide med to utseendemessig, slående like og likevel funksjonelt ulike enzymer (Kbl og BioF -se bildet). Hensikten med forsøket var å se hvorvidt det var mulig for det ene enzym A å få endret sin virkemåte i tråd med enzym B sin funksjon, i løpet av evolusjonær tidsperiode (milliarder av år). Om naturlig seleksjon virkelig førte til at svamper kunne bli spekkhoggere på mye kortere tid, og dannet mange nye proteiner underveis, burde det være mulig. Men etter nøye å ha testet mutasjoner som mest sannsynlig kunne føre til denne funksjonelle endringen, så konkluderte de at det sannsynligvis ikke var mulig, for darwinistisk evolusjon å utføre dette. (2) Tilleggsforsøk, utført av Mariclair Reeves -som i likhet med Ann Gauger var fra Biologic Institute, understøttet dette resultatet. Hun testet flittig millioner på millioner av tilfeldige mutasjoner, og søkte etter noen evolusjonær mulighet, som de andre to kunne ha oversett i det første studiet. Hun fant ingen. (3)
Som tilsvar på disse forsøkene, har de fått inn to gode spørsmål om dette resultatet, fra ikke-vitenskapsfolk. A. Det første er hvordan det overhodet er mulig å teste prosesser som tar så lang tid. Som bilde kan en estimere hvor mange år det vil ta ett ungt tre å få full størrelse, ved å måle veksten ett år og sammenligne med resten som trengs. Om en f.eks. kjenner forholdet av lodd som vinner hver uke, kan en beregne hvor lang tid det tar før en spiller vinner. Individuelle spillere vil gjøre det bedre eller verre, men i gjennomsnitt vil langtids-resultatet bli som forventet. Vitenskapelige resultater som inkluderer sjanse, som Darwins teori gjør, analyseres mye på samme måte. Til forskjell fra et lotteri derimot, så kan ikke en vitenskapelig teori appellere til flaks i håp om å slå oddsen. Tilhengere av Darwins teori må i stedet vise at noe sammenlignbart med livet i sin nåværende rikdom, er forventet utkomme når enkelt liv eksisterer.
B. Det andre gode spørsmålet er hvorvidt vitenskapsfolk som aksepterer Darwins forklaringer om livet, også kan akseptere konklusjonen til Axe og Gauger om at enzym A ikke kan endre funksjon til det som enzym B har. Mange vitenskapsfolk gjør faktisk det. Axe er faktisk i skrivende stund, ikke klar over at noen har utfordret den konklusjonen. Så kan en undres hvordan noen kan tro at naturlig seleksjon ikke er i stand til en slik liten endring, mens de påstår at den utførte så mange kjempestore. Det nåværende svaret fra evolusjonister, er at naturlig seleksjon er 'offer for sin egen suksess'. Det innebærer at naturlig seleksjon nå oppfattes som så god til å tilpasse organismer til sine omgivelser, at den nådde sluttpunkter: Organismer som er så gode til det de er, at de ikke lenger undergår evolusjonære endringer {uten tydelige miljøendringer -oversetters tilføyelse.}
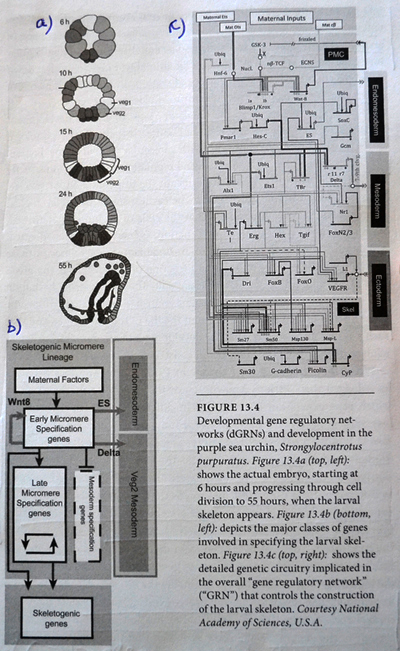 En paleontolog ved Berkeley, Charles Marshall, eksemplifiserer dette perspektivet i sin kritiske oversikt over boka 'Darwins Doubt' av Stephen C. Meyer. Meyers tese er at darwinistiske evolusjonære mekanismer ikke er i stand til å frambringe nye dyreformer, delvis fordi de ikke er i stand til å frambringe nye protein-former. Svaret Marshall antyder, er at nye dyreformer utviklet seg uten behov for nye proteinformer (6). I følge ham og andre, skjedde dette ved å 'koble om' (rewire) de regulerende gen-nettverkene (dGRNs), som kontrollerer om og når eksisterende gener slås av og på, innen et utviklende embryo. Marshall er enig i at eksperimentelle endringer av disse nettverkene vanligvis dreper embryoer, men han mener dette skulle overses, fordi "dagens dGRNs har ligget over milliarder av år med evolusjonære innovasjoner. (Mens GRNs ved tiden for dyrerekkenes oppdukking, ikke var så belemrende.)"
En paleontolog ved Berkeley, Charles Marshall, eksemplifiserer dette perspektivet i sin kritiske oversikt over boka 'Darwins Doubt' av Stephen C. Meyer. Meyers tese er at darwinistiske evolusjonære mekanismer ikke er i stand til å frambringe nye dyreformer, delvis fordi de ikke er i stand til å frambringe nye protein-former. Svaret Marshall antyder, er at nye dyreformer utviklet seg uten behov for nye proteinformer (6). I følge ham og andre, skjedde dette ved å 'koble om' (rewire) de regulerende gen-nettverkene (dGRNs), som kontrollerer om og når eksisterende gener slås av og på, innen et utviklende embryo. Marshall er enig i at eksperimentelle endringer av disse nettverkene vanligvis dreper embryoer, men han mener dette skulle overses, fordi "dagens dGRNs har ligget over milliarder av år med evolusjonære innovasjoner. (Mens GRNs ved tiden for dyrerekkenes oppdukking, ikke var så belemrende.)"
Bilde 5. Skjematisk representasjon av dGRN (bananflue) -fra 'Darwins Doubt -Fig. 13.4
Marshall og Axe er i det minste enige om at livet i sin nåværende form, motsetter seg evolusjonære endringer. De er uenige om sannsynligheten for at naturlig seleksjon noensinne har gjort noe ekstraordinært, men begge favoriserer en forklaring på livet, som virker mer hensiktsstyrt enn Darwins forklaring. Om naturlig seleksjon virkelig formet livet, så virker den mer lik en artist som former leire, enn erosjon som former sandstein. Den molekylære versjonen av det synet, er blitt hovedkritikken av konklusjonen til Axe og Gauger, samt Mariclair Reeves. Kritikere hevder nå at det var feil å forvente at enzym A skulle endre virkemåte, fordi enzymer i likhet med folk, er blitt 'perfeksjonert' til det punktet at de ikke lenger er medgjørlige i forhold til naturlig seleksjon.
Dan Tawfik (Weizmann Institute) er en talsmann for denne idéen. Han mener at 'bred-spesifikke' enzymer tjente som 'originator' for dagens spesialiserte enzymer. (7) Til sin fordel gjenkjenner Tawfik vansken med å forklare hvordan de formodede 'bred-spesifikke' enzymer oppsto i første omgang. Det måtte også ha vært skikkelige enzymer, foldede proteiner, med geometrisk komplekse aktive lokasjoner. Det er dermed uklart hva som oppnås ved å foreslå dem som forløpere. Tawfiks egen diagnose om dette, er overraskende ærlig. I følge ham har evolusjonsteorien dette 'Catch-22': Ingenting utvikler seg uten at det allerede eksisterer' (8). En kan altså ikke forvente et virksomt Y ut av noen prosess, om ikke et virksomt X gikk inn i den. Axe er enig i dette, og synes det danner håp om løsning på disputten mellom forskere. Det mest fornuftige spørsmål å stille da, er kanskje: 'hva blir igjen av en teori om opphav, så snart det medgis at den ikke forklarer hvordan organismer oppsto'?
I dette kapitlet har vi bl.a. sett at levende organismer er utsøkte helheter, så overgitt til å være det de er, at de gir et tydelig uttrykk av å være ment å være det de er. Vi skal nå se litt på det motsatte synet: Om livet ikke var ment å være det det er, så er det tilfeldig. Og av de meget få forslag til hvordan dette kunne ha skjedd, er det naturlig seleksjon som har flest meningsfeller. Da vil vi kunne vurdere naturlig seleksjon i lys av de kraftfulle linsene til 'allmenn vitenskap'.
Referanser:
1. Charles Darwin, On the Origin of Species by Means of Natural Selection, 1.utg, kap. 4, (London: John Murray, 1859, 84
2. A.K.Gauger og D. Axe, "The Evolutionary Accessibility of New Enzyme Functions: A Case Study from the Biotin Pathway," BIO-complexity, no I (2011): 1-17
3. M.A. Reeves, A.K. Gauger og D. Axe, "Enzyme Families: Shared Evolutionary History of Shared Design? A study of the GABA-Aminotrarnsferase Family," BIO-Complexity, no. 4 (2014), 1-16
6. I kap. 10 skal vi se at genomiske data motsier Marshalls forslag.
7. O. Khersonsky og D.S. Tawfik, "Enzyme Promiscuity: A mechanistic and Evolutionary Perspective," Annual Review of Biochemistry 79 (2010): 471-505
8. R. Mukhopadhyay, "Close to a Miracle: Researchers are debating the Origins of Proteins, ASBMB Today 12, no. 9 (2013): 12-13
Stoffutvalg og bilder ved Asbjørn E. Lund
{kind=link}
{kind=link}