 Livets opprinnelse, selvorganisering og informasjon
Livets opprinnelse, selvorganisering og informasjon
Brian Miller; 20. juni 2017
Oversatt herfra.
Bilde 1. Tornado kreditt: Jean Beaufort, via PublicDomainPictures.net.
I en artikkel her nylig beskrev jeg de termodynamiske utfordringene til en rent materialistisk teori for livets opprinnelse. Nå vil jeg ta opp en av de mest populære og misforståtte påstandene om at den første cellen kom fram gjennom en prosess som viste egenskapen kjent som selvorganisasjon.
 Som jeg nevnte i forrige artikkel, hevder opprinnelse-til-liv forskere ofte at livet utviklet seg i et miljø som ble drevet langt fra likevekt, ofte referert til som et ikke-likevekts-spredningssystem. I slike systemer kommer og går energi og/eller masse stadig, og denne strømmen genererer spontant "orden" som boble-mønstrene i kokende vann, trakten til en tornado eller bølgemønsteret i Belousov-Zhabotinsky-reaksjonen-her . Påstanden er at en analog type selvorganiseringsprosess kunne ha opprettet ordenen i den første cellen. Slike krav høres først rimelige ut, men de bryter helt sammen når forskjellene mellom selvorganiserende orden og cellulær orden blir undersøkt i detalj. I stedet krever livets opprinnelse komplekse cellulære maskiner og pre-eksisterende kilder til informasjon.
Som jeg nevnte i forrige artikkel, hevder opprinnelse-til-liv forskere ofte at livet utviklet seg i et miljø som ble drevet langt fra likevekt, ofte referert til som et ikke-likevekts-spredningssystem. I slike systemer kommer og går energi og/eller masse stadig, og denne strømmen genererer spontant "orden" som boble-mønstrene i kokende vann, trakten til en tornado eller bølgemønsteret i Belousov-Zhabotinsky-reaksjonen-her . Påstanden er at en analog type selvorganiseringsprosess kunne ha opprettet ordenen i den første cellen. Slike krav høres først rimelige ut, men de bryter helt sammen når forskjellene mellom selvorganiserende orden og cellulær orden blir undersøkt i detalj. I stedet krever livets opprinnelse komplekse cellulære maskiner og pre-eksisterende kilder til informasjon.
Bilde 2. Forsøk på 'selvorganisering'
Hovedårsaken til forskjellene mellom selvorganiserende og cellulær orden, er at styrende tendenser i ikke-likevektssystemer beveger seg i motsatt retning av det som trengs for både opprinnelse og vedlikehold av livet. For det første produserer alle realistiske eksperimenter om opprinnelsen til livets byggesteiner de fleste nødvendige molekyler i svært små konsentrasjoner -her, om i det hele tatt . Og de blandes sammen med 'forurensere', noe som ville hindre de neste stadiene av celledannelse. Naturen ville ha behov for spontant å konsentrere og rense livets forløpere. Den naturlige tendensen for dem ville imidlertid ha vært å diffundere og blande seg med andre kjemikalier, særlig i slike miljøer som bunnen av havet.
Konsentrasjonen av noen av livets forløpere kunne ha funnet sted i et fordampnings-basseng, men forurensningsproblemet ville da vært mye verre siden forløpere ville være mye i mindretall i forhold til 'forurensere'. Videre vil de neste stadiene av dannelse av en celle kreve at de konsentrerte kjemikalier løses opp igjen i en større vannkilde, da forskjellige forløpere måtte ha dannet seg på forskjellige steder med sterkt forskjellige startforhold. For mer informasjon om disse problemene, se her eller her.
 I tillegg kommer mange av livets byggeklosser i både høyre og venstrehånds versjoner, som er speilmotsetninger. Begge formene er produsert i alle realistiske eksperimenter i like store mengder, men livet kan bare bruke én av dem: I dagens liv, venstrehånds (L-) aminosyrer og høyrehånds sukkerarter. Livets opprinnelse ville ha krevd én form å bli stadig dominerende -her, men naturen ville drive en blanding av de to formene mot like prosenter, den motsatte retningen. Som en relatert, men mer generell utfordring, beveger alle spontane kjemiske reaksjoner nedoverbakke mot lavere fri energi. En stor del av de nødvendige reaksjonene i opprinnelsen og opprettholdelsen av livet beveger seg oppover mot høyere fri energi -her. Selv de som beveger seg nedoverbakke, går ofte for sakte til å være nyttige. Naturen ville ha måttet reversere de fleste av sine naturlige tendenser i et hvilket som helst scenario for lengre tidsperioder. Forskere har aldri observert noen slik begivenhet noensinne i universets historie.
I tillegg kommer mange av livets byggeklosser i både høyre og venstrehånds versjoner, som er speilmotsetninger. Begge formene er produsert i alle realistiske eksperimenter i like store mengder, men livet kan bare bruke én av dem: I dagens liv, venstrehånds (L-) aminosyrer og høyrehånds sukkerarter. Livets opprinnelse ville ha krevd én form å bli stadig dominerende -her, men naturen ville drive en blanding av de to formene mot like prosenter, den motsatte retningen. Som en relatert, men mer generell utfordring, beveger alle spontane kjemiske reaksjoner nedoverbakke mot lavere fri energi. En stor del av de nødvendige reaksjonene i opprinnelsen og opprettholdelsen av livet beveger seg oppover mot høyere fri energi -her. Selv de som beveger seg nedoverbakke, går ofte for sakte til å være nyttige. Naturen ville ha måttet reversere de fleste av sine naturlige tendenser i et hvilket som helst scenario for lengre tidsperioder. Forskere har aldri observert noen slik begivenhet noensinne i universets historie.
Bilde 3. Aminosyrer i to utgaver i naturen
 Disse utfordringene slått sammen bidrar til å klargjøre de dramatiske forskjellene mellom de to typene av orden:
Disse utfordringene slått sammen bidrar til å klargjøre de dramatiske forskjellene mellom de to typene av orden:
1. Selvorganiserende prosesser skaper orden (dvs. traktsky) på makroskopisk (synlig) nivå, men de genererer entropi på mikroskopisk nivå -her. I motsetning til dette krever livet at entropien på celle-størrelse format skal reduseres.
2. Selvorganiserende mønstre drives av prosesser som beveger seg mot lavere fri energi. Mange prosesser som genererer cellulær orden beveger seg mot høyere fri energi.
3. Selvorganiserings-orden er dynamisk - materialet er i bevegelse og mønstrene endres over tid. Den cellulære orden er statisk - molekyler er i faste konfigurasjoner, slik som sekvensen av nukleotider i DNA eller strukturen til cellulære maskiner.
4. Selvorganiserende orden er drevet av naturlige lover. Ordenen i celler representerer spesifiserte komplekse-molekyler -her, påtar seg svært usannsynlige ordninger som ikke er produktet av naturlige prosesser, men i stedet er arrangert for å oppnå funksjonelle mål.
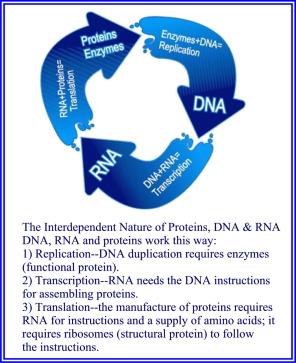
Bilde 4. Gjensidig avhengighet (dead-lock)
Disse forskjellene viser at selvorganiserings-prosesser ikke kunne ha produsert ordenen i den første cellen. I stedet krever cellulær-orden molekylære maskiner for å behandle energi fra eksterne kilder og for å lagre den i lett tilgjengelige arkiver. Og det trengte informasjon for å styre bruken av den energien mot ordentlig organisering og vedlikehold av cellen.
En enkel analogi vil demonstrere hvorfor maskiner og informasjon var avgjørende. Forskere hevder ofte at en hvilken som helst opprinnelig energikilde kunne ha gitt den nødvendige fri energi til å generere liv. Men dette kravet er som et par som kommer hjem fra en lang ferie for å finne ut at barna forlot huset i fullstendig uorden, med klær på gulvet, uvaskede tallerkener i vasken, og papirer spredt over alle arbeidsbordene. Paret hørte nylig en opprinnelse-til-livet forsker hevder at orden kunne produseres gratis fra enhver generisk energikilde. Basert på denne ideen, helles de bensin på møblene sine og setter dem i brann. De antar at energien utgått fra brannen vil organisere huset deres. Imidlertid innser de snart at ubehandlet energi skaper et enda større rot.
 Basert på denne erfaringen kjøper paret i stedet en soldrevet robot. Solceller behandler energien fra solen og konverterer den til nyttig arbeid. Men til parets skuffelse begynner roboten å kaste objekter i alle retninger. De ser nærmere på brukerhåndboken og innser at de må programmere roboten med instruksjoner for hvordan de skal utføre de ønskede oppgaver for skikkelig å rydde opp i huset.
Basert på denne erfaringen kjøper paret i stedet en soldrevet robot. Solceller behandler energien fra solen og konverterer den til nyttig arbeid. Men til parets skuffelse begynner roboten å kaste objekter i alle retninger. De ser nærmere på brukerhåndboken og innser at de må programmere roboten med instruksjoner for hvordan de skal utføre de ønskede oppgaver for skikkelig å rydde opp i huset.
Bilde 5. Eks. på frigjøring/oppbygning av energi (kredit: OpenStax Biology)
 På samme måte krevde den enkleste celle-lenke krevende maskiner, for eksempel en opprinnelig ekvivalent til ATP-syntase -her eller kloroplaster -her, for å behandle grunnleggende kjemikalier eller sollys. Det trengte også proteiner med den riktige informasjonen i deres aminosyresekvenser -her, for å folde seg til andre viktige cellulære strukturer, for eksempel portaler i cellemembranen. Og den trengte proteiner med de riktige sekvensene for å folde seg til enzymer til å drive stoffskiftet. En sentral rolle for enzymene er å koble reaksjoner som beveger seg mot lavere fri energi (f.eks. ATP -> ADP + P) til reaksjoner, for eksempel å kombinere aminosyrer i lange kjeder, som går 'oppover' -her. Energien fra den tidligere kan da brukes til å drive sistnevnte, siden netto endring i fri energi er negativ. Fri-energi barrieren er dermed overvunnet.
På samme måte krevde den enkleste celle-lenke krevende maskiner, for eksempel en opprinnelig ekvivalent til ATP-syntase -her eller kloroplaster -her, for å behandle grunnleggende kjemikalier eller sollys. Det trengte også proteiner med den riktige informasjonen i deres aminosyresekvenser -her, for å folde seg til andre viktige cellulære strukturer, for eksempel portaler i cellemembranen. Og den trengte proteiner med de riktige sekvensene for å folde seg til enzymer til å drive stoffskiftet. En sentral rolle for enzymene er å koble reaksjoner som beveger seg mot lavere fri energi (f.eks. ATP -> ADP + P) til reaksjoner, for eksempel å kombinere aminosyrer i lange kjeder, som går 'oppover' -her. Energien fra den tidligere kan da brukes til å drive sistnevnte, siden netto endring i fri energi er negativ. Fri-energi barrieren er dermed overvunnet.
Bilde 6. ATP-motorer i aktivitet
Imidlertid var energimaskiner og informasjonsrike proteiner fortsatt ikke nok. Proteiner brytes ned til slutt, og de kan ikke selvreplikere. Det var også nødvendig med ekstra maskiner for å kontinuerlig å produsere nye protein-utskiftninger. Proteinenes sekvens-informasjon måtte også ha blitt lagret i DNA ved bruk av en genetisk kode, der hver aminosyre ble representert ved en serie av tre nukleotider kjent som et kodon på samme måte som bokstaver er representert i morse-koden med prikker og streker. Imidlertid eksisterer ingen identifiserbar fysisk forbindelse -her mellom individuelle aminosyrer og deres respektive kodoner. Spesielt er ingen aminosyrer (f.eks. Valin) mye sterkere tiltrukket av ett bestemt kodon (f.eks. GTT) enn til noen andre. Uten en slik fysisk forbindelse kunne ikke en rent materialistisk prosess troverdig forklare hvordan aminosyre-sekvensene ble kodet inn i DNA. Derfor må samme informasjon i proteiner og i DNA være kodet separat.
Bilde 7. Konklusjon ang.livets opprinnelse

I tillegg dekodes informasjonen i DNA tilbake til proteiner -her, ved bruk av ribosomer, tRNAer og spesielle enzymer kalt aminoacyl-tRNA-sytetaser -her (aaRS). AaRS-ene binder de riktige aminosyrene til de korrekte tRNAene som er knyttet til de riktige kodonene, slik at disse enzymer inneholder dekodings-nøkkelen i deres 3D-struktur. Alt liv bruker samme prosess, så den første cellen fungerte nesten helt sikkert på samme måte. Imidlertid kan det ikke eksistere noen mulig sammenheng mellom kodings og dekodings-prosessene, siden aaRSs strukturer er et resultat av deres aminosyresekvenser, som tilfeldigvis er en del av informasjonen som er kodet i DNA. Derfor måtte dekoding ha utviklet seg uavhengig av kodingen, men de måtte bruke samme kode. Og de måtte oppstå samtidig, siden hver av dem er ubrukelig uten den andre.
Alle disse fakta indikerer at koden og sekvensinformasjonen i proteiner/DNA eksisterte forut for den opprinnelige cellen. Og det eneste stedet de kunne eksistere utenfor et fysisk medium er i et sinn, som peker på design.
Stoffutvalg og bilder ved Asbjørn E. Lund