 Genetikk og makroevolusjon
Genetikk og makroevolusjon
(fra kap. 2 i ‘The Design of Life’, W. Dembski og J. Wells)
Bilde 1. Utgangspunkt
I 'Origin of Species' kjempet Darwin for at ingen eksisterende art var individuelt skapt. I stedet hevdet han at hver art nedstammet fra pre-eksisterende arter, som et resultat av naturlig seleksjon. Han argumenterte at alle arter sporer sitt opphav tilbake til én eller noen få originale elementer eller former. I følge hans teori utviste organismer nye trekk når naturlig seleksjon 'velger' blant tallrike naturlige genetiske varianter. Da hevder teorien at visse varianter vil bli overført til fremtidige generasjoner, om de gir sin besitter et 'konkurransefortrinn' fremfor andre organismer i populasjonen, og derfor gir flere etterkommere. Darwin argumenterte at som evolusjonen gikk framover, ville fremgangsrike nye trekk akkumuleres, inntil en ny art ble dannet.
Mendel skilte dominante fra recessive gener. I hans opprinnelige eksperiment med erter, er begge former av gener (kjent som alleler) tilstede, men bare én blir uttrykt for et gitt trekk. Men ikke alle trekk blir assosiert med et enkelt gen. Noen, f.eks. hudfarge, blir assosiert med mange gener som arbeider i kombinasjon med hverandre. Fargekombinasjoner hos avkommet, vil da kunne variere over hele spekteret av fargekombinasjoner, der ytterpunktene er mest sjeldne. Likevel taper ikke gener sin identitet ved å kombineres, med gir ulike resultat ved å bli kombinert på ulike måter. Om det er en fordel med mørk hudfarge, vil naturlig seleksjon foretrekke dette, slik at kombinasjon av gener som produserer mørk hudfarge vil bli etablert i denne regionen. Men det ville ikke dermed bli en endring fra en art til en annen. Endringen skjer bare i utbredelsen, frekvens-forekomsten av visse gen-kombinasjoner.
 Genetisk variasjon innebærer i seg selv en fordel for en populasjon. Men nye kombinasjoner kan i utgangspunktet like gjerne skyldes kombinasjoner av eksisterende gener, som viste seg fordelaktig, ikke nødvendigvis opprinnelsen av nye gener. I eks. med spurv som gjennomgår endringer, så trenger ikke gen-forandring være årsaken. Men gener kan 'skyfles' omkring, og frambringe nye kombinasjoner av gener som allerede tidligere fantes. Selv i relativt små populasjoner, kan det være større potensiell diversitet i kroppsform, enn det syntes å være i begynnelsen. En eskimo fra Alaska og en afrikaner fra Nil-regionen tilhører samme art, men kontrasten i kroppsform er likefullt spektakulær. Afrikanerens lange armer og ben er en fordel i varme soner, men vil være en ulempe for eskimoen. Det er kombinasjon av fikserte gener, ikke selve de individuelle genene, som i dette tilfellet utsettes for naturlig seleksjon. Om en populasjon ikke er i stand til å utvide seg til nye områder, eller tilpasse seg varierende forhold, så vil den antageligvis forbli liten eller dø ut.
Genetisk variasjon innebærer i seg selv en fordel for en populasjon. Men nye kombinasjoner kan i utgangspunktet like gjerne skyldes kombinasjoner av eksisterende gener, som viste seg fordelaktig, ikke nødvendigvis opprinnelsen av nye gener. I eks. med spurv som gjennomgår endringer, så trenger ikke gen-forandring være årsaken. Men gener kan 'skyfles' omkring, og frambringe nye kombinasjoner av gener som allerede tidligere fantes. Selv i relativt små populasjoner, kan det være større potensiell diversitet i kroppsform, enn det syntes å være i begynnelsen. En eskimo fra Alaska og en afrikaner fra Nil-regionen tilhører samme art, men kontrasten i kroppsform er likefullt spektakulær. Afrikanerens lange armer og ben er en fordel i varme soner, men vil være en ulempe for eskimoen. Det er kombinasjon av fikserte gener, ikke selve de individuelle genene, som i dette tilfellet utsettes for naturlig seleksjon. Om en populasjon ikke er i stand til å utvide seg til nye områder, eller tilpasse seg varierende forhold, så vil den antageligvis forbli liten eller dø ut.
Bilde 2. 50 protein-former pr. gen
Molekylær basis for gener og evolusjon
Fordi sekvenser av DNA er i stand til å kode for proteiner, så er det presist å beskrive slike kodende sekvenser av DNA som beskjeder. Faktisk er det en dyp sammenheng mellom nukleotid-sekvenser i DNA som koder for proteiner og sekvenser av alfanumeriske tegn, som overbringer beskjeder. Tidligere celle-biolog ved Universitetet i London, E.J. Ambrose, uttrykte dette slik: "Det er en beskjed, hvis rekkefølgen av baser i DNA kan bli oversatt av cellen til en vital aktivitet som er nødvendig for overlevelse eller reproduksjon." (10) Bare én ekstremt liten andel av det astronomiske antall alfanumeriske sekvenser i dette skjermbildet, kan inneholde samme mening som denne. Således er det en ekstremt liten andel av det astronomiske antall DNA-sekvenser, som kan kode for et funksjonelt protein av en gitt type. Selv om ikke Cambridge paleontolog Simon Conway Morris er noen fortaler for Intelligent Design (ID), så gjenkjenner han dette faktum og dets betydning. 'Isolerte øyer' forsyner havner av biologiske muligheter i et hav av mistilpasning. Ikke til å undres at argumentet for design og intelligent planlegging har en slik varig appell." (11)
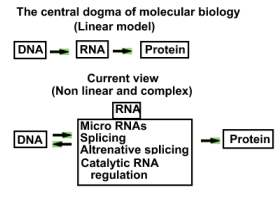 Like fullt mener neo-darwinister at de har svaret på DNA's språklignende evne til å kode for biologisk signifikante strukturer, et svar som ikke innbefatter ID. Deres svar går (som kjent) på at naturlig seleksjon virker på gen-mutasjoner. Mutasjoner er molekylære endringer i DNA, som grupperes i to klasser. A) Punkt-mutasjoner, Her opptrer endringene i individuelle nukleotide- baser i DNA. (12) B) Kromosom-mutasjoner, som involverer hele seksjoner av DNA. Kromosom-muasjoner kan resultere i duplikat av et DNA-segment, tapet av det, eller rekombinasjon på et annet sted for samme eller et et ulikt DNA-molekyl, eller endog inversjonen innen dette molekylet. Punkt-mutasjoner er sjeldne. Det har vært kjent en stund at et gen i gjennomsnitt endrer seg bare 1: 100.000 eller 1:1.000.000 reproduksjoner. (13) En annen måte å utrykke punkt-mutasjons raten, er hvor mang sædceller som inneholder minst én mutasjon. Studier viser at et mutant gen i gjennomsnitt vil inntreffe for hver 10. til 100. sædcelle. Mens en ikke kjenner underliggende årsak for denne grunnleggende punkt-muasjosjons raten, så kan raten økes ved omgivelsesfaktorer som varme, kjemikalier eller stråling.
Like fullt mener neo-darwinister at de har svaret på DNA's språklignende evne til å kode for biologisk signifikante strukturer, et svar som ikke innbefatter ID. Deres svar går (som kjent) på at naturlig seleksjon virker på gen-mutasjoner. Mutasjoner er molekylære endringer i DNA, som grupperes i to klasser. A) Punkt-mutasjoner, Her opptrer endringene i individuelle nukleotide- baser i DNA. (12) B) Kromosom-mutasjoner, som involverer hele seksjoner av DNA. Kromosom-muasjoner kan resultere i duplikat av et DNA-segment, tapet av det, eller rekombinasjon på et annet sted for samme eller et et ulikt DNA-molekyl, eller endog inversjonen innen dette molekylet. Punkt-mutasjoner er sjeldne. Det har vært kjent en stund at et gen i gjennomsnitt endrer seg bare 1: 100.000 eller 1:1.000.000 reproduksjoner. (13) En annen måte å utrykke punkt-mutasjons raten, er hvor mang sædceller som inneholder minst én mutasjon. Studier viser at et mutant gen i gjennomsnitt vil inntreffe for hver 10. til 100. sædcelle. Mens en ikke kjenner underliggende årsak for denne grunnleggende punkt-muasjosjons raten, så kan raten økes ved omgivelsesfaktorer som varme, kjemikalier eller stråling.
Bilde 3. Fra lineær til ikke-lineær modell
Liksom ikke lesbarheten økes ved at bokstaver tilfeldig skiftes om, som kan skje her, så har mutasjoner samme effekt i den biologiske verden. Med unntak av ekstreme forhold der harde omgivelser utøver enormt påtrykk for seleksjon, som når bakterier utsatt for antibiotika utvikler resistens, så er ingen tjenlige punktmutasjoner kjent. Slike tjenlige punktmutasjoner involverer små-skala endringer i enkelte protein-molekyler og forsyner ikke bevis for makroevolusjon. Enn videre når omgivelses-trykket blir redusert, synes den angitte fordel å gå tapt. F.eks. når antibiotika fjernes fra bakterie-kulturen, så dukker den opprinnelige bakterien opp igjen og dominerer populasjonen. Dette er ikke noen fullskala-evolusjon. Det er ett steg fram og ett tilbake.
 På grunn av slike begrensninger på punkt-mutasjoner ser noen neo-darwinister til kromosom-mutasjoner for å få fram makro-evolusjonære endringer. De betrakter gen-duplisering som spesielt viktig i denne sammenheng. Det er fordi når ett gen er duplisert, så er det potensielt tilgjengelig for å tjene en annen funksjon. Normalt dedikeres ett gen til en eksisterende funksjon, men om det dupliseres, så det dupliserte genet overflødig, og kreves dermed ikke for å tjene eksisterende funksjon, og dermed fri fra seleksjonspress for å utøve den funksjonen. I følge neo-darwinister, er det dupliserte genet således sannsynlig å oppnå en ny funksjon, når det navigerer gjennom det 'genetiske rommet', som så kan forsyne basis for makroevolusjonær endring.
På grunn av slike begrensninger på punkt-mutasjoner ser noen neo-darwinister til kromosom-mutasjoner for å få fram makro-evolusjonære endringer. De betrakter gen-duplisering som spesielt viktig i denne sammenheng. Det er fordi når ett gen er duplisert, så er det potensielt tilgjengelig for å tjene en annen funksjon. Normalt dedikeres ett gen til en eksisterende funksjon, men om det dupliseres, så det dupliserte genet overflødig, og kreves dermed ikke for å tjene eksisterende funksjon, og dermed fri fra seleksjonspress for å utøve den funksjonen. I følge neo-darwinister, er det dupliserte genet således sannsynlig å oppnå en ny funksjon, når det navigerer gjennom det 'genetiske rommet', som så kan forsyne basis for makroevolusjonær endring.
Bilde 4. Punkt-mutasjoner
Selv om denne historien har en viss troverdighet, så faller den raskt fra hverandre når den testes. (14) For det første er det bare avgjort av tilfeldigheter hvordan det dupliserte genet endres. Punkt-mutasjoner er som nevnt meget sjeldne, og kromosom-mutasjoner på et duplisert gen innebærer videre handlinger ved tilfeldighet som re-arrangerer og reposisjonerer genet. Men kanskje mutasjoner kan forsyne råmaterialet for makroevolusjon, når den blir passende renset via naturlig seleksjon.
La oss se hva slags genetiske endringer makroevolusjon krever. Da må en først se hvilke nøkkeltrekk ved biologiske organismer, som makroevolusjon må forklare. En organismes evne til å fungere tilfredsstillende, krever en hel pakke med tilpasninger, d.v.s. ett sett strukturer som er nøye koordinert med hverandre, for å hjelpe organismen å leve. Utfordringen for makroevolusjon, er å fremskaffe slike tilpasnings-pakker. Her kan vi betrakte giraffen, som Darwin skrev om i 'Origin of Species': "Giraffen .. opptar føde utenfor rekkevidde av .. andre hovdyr som bebor samme område, og dette må være en stor fordel for den..." (15)
 Imidlertid er ikke fordelen av giraffens lange hals, for å 'browse' på de høyere grenene på trærne så åpenbar som Darwin får det til. F.eks. er hunnene to fot kortere. Om en så lang nakke, som hannens, virkelig var nødvendig, ville hunene snart dø av sult, og giraffen ville dø ut. Darwin hadde rett i at giraffen var 'skjønt tilpasset', men han hadde ikke tilstrekkelig informasjon til å skjønne hele omfanget og raffinementet i tilpasningene. Observer noen giraffer som spiser og drikker i zoologisk-have, og du vil se at de ikke bare strekker hodet for å spise de øverste grenene, men også bøyer seg ned til bakken for å spise gress og drikke vann. Gitt sine lange bein, så trenger giraffer en lang hals, mindre for å rekke opp, enn å rekke ned til bakken.
Imidlertid er ikke fordelen av giraffens lange hals, for å 'browse' på de høyere grenene på trærne så åpenbar som Darwin får det til. F.eks. er hunnene to fot kortere. Om en så lang nakke, som hannens, virkelig var nødvendig, ville hunene snart dø av sult, og giraffen ville dø ut. Darwin hadde rett i at giraffen var 'skjønt tilpasset', men han hadde ikke tilstrekkelig informasjon til å skjønne hele omfanget og raffinementet i tilpasningene. Observer noen giraffer som spiser og drikker i zoologisk-have, og du vil se at de ikke bare strekker hodet for å spise de øverste grenene, men også bøyer seg ned til bakken for å spise gress og drikke vann. Gitt sine lange bein, så trenger giraffer en lang hals, mindre for å rekke opp, enn å rekke ned til bakken.
Giraffen er en integrert tilpasset 'pakke', hvis deler er nøye koordinert med hverandre. For å passe suksessfylt i sin omgivelses-nisje, trengte giraffen lange bein. Men da trengte den også en lang hals, og for å kunne benytte denne lange halsen så var ytterligere tilpasninger nødvendig. Forskjellen i blodtrykk øverst og nederst i halsen hos giraffen er betydelig. Blodtrykket som genereres i hjertet, må være formidabelt for å nå opp til hodet. Men da oppstår en potensielt farlig situasjon, når den bøyer hodet mellom frambeina. Blodtrykk sammen med blodets vekt, kunne produsere så mye trykk i hodet, at uten sikkerhetsanordninger, ville blodårene springe.
Bilde 5. En giraff utgjør ei helhetlig 'pakke'
Slike sikkerhetsanordninger er imidlertid på plass. Giraffens tilpasningspakke inkluderer et koordinert system av blodtrykks-kontroller. Trykkmålere langs nakke-arteriene måler trykket, og kan signalere aktivering av andre mekanismer for å imøtekomme trykk-økninger i det giraffen drikker eller gresser. Sammentrekning av arterie-veggene, evnen til å skubbe blod som gjennomstrømmer hjernen, og en liten vev av små blodårer mellom arteriene og hjernen kontrollerer blodtrykk i giraffens hode. Giraffens tilpasninger skjer ikke i isolasjon, men må alle være nøye koordinert i en enkel, høyst spesifisert organisme. Problemet med integrerte pakker, er at de ikke tilbyr noen fordel inntil hele pakken er på plass. (16) Denne inter-avhengigheten av strukturer foreslår sterkt en top-down design, som er i stand til å forvente de totale ingeniør-kravene for organismer lik giraffen.
For å utvikle nye tilpasnings-pakker møter populasjoner derfor en informasjons-hindring. Én måte å se denne hindringen, er i fenomenet med fylogenetisk treghet. Det betegner en populasjons tendens til å vedlikehold en gjennomsnitts morfologi så vel som en begrenset variabilitet, omkring populasjonens gjennomsnitt. Hvordan kan mutasjoner klare å beseire slike hindringer? Det er ikke klart at de kan det. Kromosom-mutasjoner kan utveksle deler av gen-sekvenser, men det er ikke bevis for slik 'nye' gener kan forsyne den stadige akkumulering av nye trekk, som darwinistisk evolusjon trenger for å være effektiv. Kromosom-mutasjon skyfler bare omkring på eksisterende gener.
Den eneste kjente måte å introdusere genuint ny informasjon, er ved mutasjoner som endrer nukleotid-basene for individuelle gener. Det er ulikt kromosom-mutasjoner, der seksjoner av DNA blir duplisert, invertert, tapt eller flyttet i DNA-molekylet. Punkt-mutasjoner, ikke bare re-arrangerer, men endrer fundamentalt strukturen til eksisterende gener. Slike mutasjoner resulterer typisk fra tilfeldige kopierings feil ved DNA og blir intensivert gjennom å bli utsatt for hete, kjemikalier eller stråling. Kan kromosom og punkt -mutasjoner som virker sammen, forsyne råmaterialet for makroevolusjonære endringer? Som primærkilde for evolusjonære nyheter i Neo-Darwinistisk (ND) teori, er mutasjoner blitt studert intensivt gjennom det siste halve århundre.
 Frukt-flua (Drosophila) er et slikt tilfelle, som er lett manipulerbar og har kort generasjonstid. Den har blitt utsatt for tallrike eksperimenter, ved å bombardere den med stråling, har forskere nå en rimelig klar idé om hva slags mutasjoner som kan inntreffe. Det er ikke bevis for at mutasjoner kan produser nye fungerende strukturer. F.eks. har mutasjoner produsert forkrøblede, forstørrede - og forminskede vinger. Men de har ikke produsert en ny type fungerende vinge. For å oppsummere, så må det genereres en tilpasnings-pakke som krever integrerte, systematiske endringer. Videre må kilden til slike endringer tildele massive mengder av ny fungerende informasjon til en organisme. Slik informasjon har en ikke noe bevis for oppstår ved mutasjon og seleksjon. Faktisk gir den ikke noe bevis for å være reduserbar til materie og energi i det hele. Norbert Wiener, en av grunnleggerne av informasjons-teori bemerker: "Informasjon er informasjon, ikke materie eller energi. Ingen materialisme som ikke innrømmer dette, kan overleve nåtiden." (18) På samme vis som informasjon på dette skjermbildet, er ulikt bits og pixler, så er informasjonen i biologiske systemer forskjellig fra sine materielle bestanddeler. Hva er informasjonskilden til å bygge en tilpasset pakke? Som med informasjon i skrevne beskjeder og ingeniør-systemer, så er eneste kjente kilde som er kjent for å være i stand til å generere informasjon, slik som vi ser i biologien, intelligens.
Frukt-flua (Drosophila) er et slikt tilfelle, som er lett manipulerbar og har kort generasjonstid. Den har blitt utsatt for tallrike eksperimenter, ved å bombardere den med stråling, har forskere nå en rimelig klar idé om hva slags mutasjoner som kan inntreffe. Det er ikke bevis for at mutasjoner kan produser nye fungerende strukturer. F.eks. har mutasjoner produsert forkrøblede, forstørrede - og forminskede vinger. Men de har ikke produsert en ny type fungerende vinge. For å oppsummere, så må det genereres en tilpasnings-pakke som krever integrerte, systematiske endringer. Videre må kilden til slike endringer tildele massive mengder av ny fungerende informasjon til en organisme. Slik informasjon har en ikke noe bevis for oppstår ved mutasjon og seleksjon. Faktisk gir den ikke noe bevis for å være reduserbar til materie og energi i det hele. Norbert Wiener, en av grunnleggerne av informasjons-teori bemerker: "Informasjon er informasjon, ikke materie eller energi. Ingen materialisme som ikke innrømmer dette, kan overleve nåtiden." (18) På samme vis som informasjon på dette skjermbildet, er ulikt bits og pixler, så er informasjonen i biologiske systemer forskjellig fra sine materielle bestanddeler. Hva er informasjonskilden til å bygge en tilpasset pakke? Som med informasjon i skrevne beskjeder og ingeniør-systemer, så er eneste kjente kilde som er kjent for å være i stand til å generere informasjon, slik som vi ser i biologien, intelligens.
Bilde 6. Drosophila -mye utforsket
Hvor mange gener må endres?
Nevnte celle-biolog E.J. Ambrose estimerte at det er "høyst usannsynlig at færre enn fem gener kunne være involvert i selv den enkleste nye struktur i en organisme." (19) Ambrose registrerte at ikke mer enn 1 ikke-skadelig mutasjon oppsto pr. generasjon i en populasjon på 1000 stk. De fleste gener har en mutasjons-frekvens mindre enn 1: 100.000 og de fleste av de mutasjonene er skadelige. Forutsetter en uavhengighet, blir dermed sannsynligheten for at to mutasjoner skal inntreffe: 1: 1.000.000 (tusen ganger tusen). Sannsynligheten for fem ikke-skadelige mutasjoner, blir dermed 1: (10 opphøyd i 15). For alle praktiske formål er det ingen sjanse for at disse fem mutasjonene kunne inntreffe i løpet av livssyklusen til en enkelt  organisme.
organisme.
Men anta at fem ikke-skadelige mutasjoner oppsto i gen-poolen til en enkelt art (i populasjonen heller enn en enkelt organisme). Anta videre at disse mutasjonen inntraff over tid, og ble bevart i hetero-zygot tilstand. Gitt rekombinasjonspotensialet fra omfattende avl, kunne ikke de fem genene evt. komme sammen i en etterfølger? 'Hardy-Winberg' loven hevder at i fravær av seleksjon, eller andre utenforliggende krefter, så fører tilfeldig parring til at proporsjonen av gener forblir de samme i en populasjon fra en generasjon til neste. Følgelig vil proporsjonen av disse fem muterte genene i forhold til deres ikke-muterte motparter, forbli konstant i artens populasjon.
Bilde 7. Mutert hox-gen i bananflue
Hva er sjansen for at individuelle organismer som bærer disse separate genene, ville finne hverandre over en populasjon på, f.eks. en million? Organismene ville trenge å pare seg med hverandre i rett tid og rett rekkefølge, for å kombinere alle fem gener i et enkelt individ. Videre måtte det resulterende sett av nye gener kode for en helt ny struktur for å øke organismens nivå av kompleksitet over det til sine foreldre. Men et slikt scenario synes ytterligere usannsynlig, i det det krever en opphopning av sammentreff bygd på sammentreff.
Selv om en slik forklaring var mulig, er det enda mer å forklare. Anta at vårt sett på fem gener ble fiksert i ett individuelt kromosom, der de ikke blir uttrykt i hetero-zygot tilstand, fordi de komplementære genene til de matchende kromosomene er dominante. Om de mot formodning, skulle være lokalisert i samme region av kromosomet, så måtte en ytterligere mutasjon i et annet gen formodentlig endre den regionen fra recessivt til dominant (virke som en gen-svitsj for gen-clusteret). Kunne en slik konvergens av gener i et kromosom, frembringe formasjonen til en ny struktur, i en individuell organisme? Slike fem-eller seks gens clustere er nå kjent å kontrollere vinge-farge til sommerfugler (Papilio dardanus) i Afrika. (20) Men å kontrollere fargedannelser når ikke opp til å danne nye komplekse strukturer. Ingen eks. med fem-seks gens clustere er til nå kjent, for å kunne redegjøre for komplekse strukturer.
Ambrose la også merke til at selv den enkleste biologiske struktur ville kreve mange flere enn de fem genene han betraktet. Han påpekte også usannsynligheten ved å få de rette genene i ett cluster, blekner til ubetydelighet når vi erkjenner at det må være en tett funksjonell-integrasjon mellom de individuelle genene i ett cluster, som også må være integrert i hele organismen. Ambrose konkluderte derfor at "hypoteser om opprinnelsen til arter faller til jorden, om det ikke er akseptert at et intensivt input av ny informasjon blir introdusert ved isolasjons-tidspunktet til det nye parende-par." (22)
 Evo-devo
Evo-devo
I slutten av dette kapitlet vender vi oss til et nyere forslag av darwinister for å gjøre rede for makro-evolusjon, nemlig evo-devo (evolutionary development biology). Evo-devo forsøker å blande de to underdisipliner i biologi: evolusjons-biologi, som studerer mekanismene som endrer populasjoner av organismer over generasjoner, og utviklingsbiologi som studerer hvordan individuelle organismer vokser fra unnfangelse til modenhet. Evo-devo tar som utgangspunkt at genetiske mekanismer er nøkkelen både til evolusjons og utviklingsbiologi. Blandingen av disse, ser derfor på genene som influerer utvikling og også i prinsippet kunne influere endringer i utvikling og således lede til makroevolusjonære endringer.
Hva f.eks. om et gen som kontrollerer utvikling endrer seg tidlig i utviklingen? Endog en liten endring kunne ha store konsekvenser for organismens anatomi og fysiologi. Tenk på en pil siktet inn mot et mål. Om en endret bane til pila i starten, så ville den treffe langt unna målet. På det viser søker evo-devo å ta en vei utenom den tradisjonelle neo-darwinistiske i forhold til makro-evolusjon. I kontrast til stødig akkumulering av små endringer, lover evo-devo raske evolusjonære endringer til liten kostnad, nemlig ved å mutere noen få nøkkel-gener, som kontrollerer tidlig utvikling.
Bilde 8. Genetiske mutasjoner på bananflue
Men til tross for lovende start-løfter, synes evo-devo å ha kommet til en tistand av krise. Dens studie av gener fortsetter i et raskt tempo, og feltet gjør visse fremskritt i hvordan genetiske utviklingsmekanismer assisterer mikroevolusjonære endringer (sommerfuglers farge-flekker). Men W. Jeffrey, en evolusjonsbiolog ved University of Maryland, konkluderer at "evo-devo's forsøk å forstå hvordan genetiske utviklings-mekanismer induserer makro-evolusjonære endringer, er i en blindvei." (24) Problemet er at evo-devo ser på konserverte gener, som er hovedsakelig de samme over vidt forskjellige organismer, ofte i ulike dyre-rekker, for å studere hvordan makro-evolusjonære endringer kunne ha inntruffet. Men det reiser fundementale problemer. El. Pennisi erklærer i en 2002 rapport om evo-devo i Science: "Listen [om konserverte gener] gir ingen innsikt i hvordan organismer med de samme gener til slutt kom til å bli så ulike." (25)
For å forstå problemet, la oss ta et trinn tilbake. For ca. 125 år siden observerte biologer at noen deler av noen dyr av og til utviklet seg lik andre kroppsdeler, som normalt befant seg andre steder. F.eks. utviklet et insekts antenne seg i likhet med et ben, se Bilde 7. Slike endringer ble kalt homeotiske ('blir like') av W. Bateson i 1894 for å indikere at berørt del er 'blitt lik' en annen del av organismen. (26) Med starten av moderne genetikk, ble slike endringer sporet til mutasjoner i 'homeotiske gener' som spesifiserer identitet til visse grupper av celler i løpet av embryotisk utvikling. Utgjør så homeotiske gener et nøkkel-bevis for makroevolusjon? Simpel logikk viser at svaret er: Nei. (27) Nettopp fordi homeotiske gener er universelle, så kan de ikke forklare forskjeller i organismer grunnet makroevolusjonære endringer. Her er grunnen: Dersom biologiske strukturer blir bestemt av sine gener, slik Neo-Darwinisme (ND) forutsetter, så må ulike strukturer bestemmes av ulike gener. Om de samme gener er implisert i strukturer så radikalt forskjellige som en bananflues ben og hjernen til ei mus, eller et insekts øye og øyet til mennesker og blekksprut, så bestemmer ikke genet særlig mye av noe som helst.
På lignende måte i å fortelle oss hvordan et embryo styrer sine celler til en av atskillige innebygde utviklingsstier, forteller ikke homeotiske gener oss noe om hvordan de aktuelle biologiske strukturene blir formet. I det homeotiske gener viser seg å bli mer og mer universelle, så viser kontrollen de utøver i utviklingen viser seg å bli mindre og mindre spesifikk. For å oppsummere har utviklings genetikere funnet at gener som synes å være mest viktige i utviklingen er merkelig like i mange ulike typer av dyr. I starten ble det sett som bevis for genetiske program som styrte utviklingen. Men biologer innser nå at dette utgjør et paradoks: om gener styrer utvikling, hvorfor produserer like gener så ulike dyr? Hvorfor blir en larve til en sommerfugl i stedet for til en sel?
Til slutt er problemet som utviklings genetikere har funnet, at endringene som trengs i utviklingen for å iverksette makro-evolusjonære endringer, simpelthen ikke inntreffer. Et utviklingsembyo tar, i motsetning til en pil, kraftfulle trinn for å komme tilbake på kurs og rekke sitt utviklings endemål, uansett hvor tidlige i utviklingen den blir forstyrret. Utviklings-forløp er selv-korrigerende på måter som pilbaner ikke er.
 Utviklingsbiologer studerer ofte embryogenese ved å forstyrre embryo-utvikling. Bemerkelsesverdig er at selv om forstyrrelser iverksetter deformiteter, så blir sluttmålet for utviklingen ikke endret. Om de overlever, blir bananfluer alltid bananfluer, froskeegg blir alltid frosker, og museegg blir mus. Ikke engang arten endrer seg. Ethvert embryo er på et vis programmert til å utvikle seg til en spesiell dyreart.
Utviklingsbiologer studerer ofte embryogenese ved å forstyrre embryo-utvikling. Bemerkelsesverdig er at selv om forstyrrelser iverksetter deformiteter, så blir sluttmålet for utviklingen ikke endret. Om de overlever, blir bananfluer alltid bananfluer, froskeegg blir alltid frosker, og museegg blir mus. Ikke engang arten endrer seg. Ethvert embryo er på et vis programmert til å utvikle seg til en spesiell dyreart.
Bilde 9. Forutsetninger bestemmer ofte resultat
Verdt å merke er at slike program ikke er genetiske program. Visst spiller gener en rolle i utviklingen, men å si at de kontrollerer eller bestemmer utviklingen er en stor overdrivelse. F.eks. når et eggs gener er fjernet og erstattet med gener fra en annen type dyr, følger utviklingen programmet til det opprinnelige egget, inntil embryoet dør av mangel på rette proteiner. (Den sjeldne unntakelsen involverer dyr som normalt kunne pare seg til å produsere hybrider.) Fremgangsmåten i 'Juraissic Park' med å injisere dinosaur-gener i strutse-egg, for å produsere en Tyrannosaurus rex, utgjør spennende fiksjon, men den ignorerer vitenskapelige fakta.
Hva med å mutere DNA i stedet for å erstatte den fullstendig? Ved å benytte en teknikk som kalles 'saturasjon-mutagenese' har biologer funnet at mutasjoner i utviklings gener, ofte fører til død eller deformitet, men de har aldri produsert anatomiske endringer, som tjener organismen. Videre endrer aldri DNA-mutasjoner endemålet for embryonisk utvikling. De kan ikke engang endre arten. Et embryo trenger de rette genene for å lage nye proteiner, og dets utvikling lider uten dem. Men å være avhengig er forskjellig fra å være styrt/kontrollert av dem. Et hus under konstruksjon trenger bygningsmaterialer, men disse materialene bestemmer ikke dens grunnplan.
Om DNA var i kontrollen av utviklingen, skulle en være i stand til å produsere en kopi av seg selv, ved å injisere sitt DNA i et menneske-egg, som hadde sitt eget DNA fjernet. Men en slik klone ville ikke være en identisk (yngre)  kopi av deg selv. Hvordan det ser ut ville i stor grad være avhengig av informasjon i egget uten kjerne, som mottok ditt DNA. Ikke engang eneggede tvillinger er identiske kopier av hverandre. De skiller seg ofte i fysiske karakteristika, og alltid i temperament og interesser. Likevel deler 'identiske' tvillinger ikke bare samme DNA, men også samme egg, og vanligvis samme livmor. Endog ikke-perfekt likhet av eneggede tvillinger krever mer enn samme DNA.
kopi av deg selv. Hvordan det ser ut ville i stor grad være avhengig av informasjon i egget uten kjerne, som mottok ditt DNA. Ikke engang eneggede tvillinger er identiske kopier av hverandre. De skiller seg ofte i fysiske karakteristika, og alltid i temperament og interesser. Likevel deler 'identiske' tvillinger ikke bare samme DNA, men også samme egg, og vanligvis samme livmor. Endog ikke-perfekt likhet av eneggede tvillinger krever mer enn samme DNA.
Bilde 10. Utvikling av celle -tidlig stadium
Et annet bevis mot at DNA kontrollerer utviklingen, i motsetning til bare å influere den, -som det visselig gjør, kommer fra det faktum at voksne celler inneholder samme DNA som det befruktede egg de stammer fra. Likevel skiller cellene i et voksent individ seg merkbart fra hverandre i form og funksjon. Men hvis de har samme DNA, hvorfor er de så ulike? Del av svaret er at hver celletype, som styres av ulike epigenom, benytter bare en del av sitt genetiske repertoar, ved at faktorer utenfor DNA slår på passende gener. Men om utvikling krever at DNA blir styrt av faktorer utenom seg selv, så kontrollerer ikke DNA utviklingen.
Hvorfor fortsetter da synet at genetiske program kontrollerer utviklingen å være så populært? Svaret avhenger i stor grad med dets logiske sammenheng med ND. Genetiske program er et korollar av ND-syntese av Mendels genetikk og Darwinistisk evolusjon. I følge ND forsyner genetiske muasjoner råmaterialet for evolusjon og naturlig seleksjon modifiserer organismer gjennom endringer i gen-frekvenser. Utvikling er hva som styrer ei enkelt celle til å bli ei mus i stedet for en orm. Herunder, dersom evolusjon kan endre ormer til mus ved å modifisere deres gener, så må det skje ved å modifisere gener som styrer utviklingen. Motsatt om utviklingen blir styrt av noe annet enn gener, så må evolusjon skyldes noe annet enn genetiske mutasjoner og endringer i gen-frekvenser. Om forestillingen at genetiske program styrer utviklingen er feilaktig, så er også ND det. ND fastsetter logisk, kontroll av embryo-utviklingen til genetiske program.
 Universaliteten av homeotiske gener i embryo-utvikling er antatt å skyldes deres nærvær i en felles stamfar, men fraværet av bevis viser at den 'felles stamfaren' mangler trekkene som disse homeotiske genene formodentlig besitter. Dette er et seriøst problem, sett fra ND side, da komplekse gener i følge dem gradvis utvikler ved å tilføre selektive fordeler til organismer som besitter dem. Men gen-sekvenser overdrar selektive fordeler bare om de programmerer utviklingen av nyttige tilpasninger. Om et primitivt dyr besatt homeotiske gener, men manglet alle tilpasninger som nå assosieres med dem: Hvordan utviklet da homeotiske gener seg?
Universaliteten av homeotiske gener i embryo-utvikling er antatt å skyldes deres nærvær i en felles stamfar, men fraværet av bevis viser at den 'felles stamfaren' mangler trekkene som disse homeotiske genene formodentlig besitter. Dette er et seriøst problem, sett fra ND side, da komplekse gener i følge dem gradvis utvikler ved å tilføre selektive fordeler til organismer som besitter dem. Men gen-sekvenser overdrar selektive fordeler bare om de programmerer utviklingen av nyttige tilpasninger. Om et primitivt dyr besatt homeotiske gener, men manglet alle tilpasninger som nå assosieres med dem: Hvordan utviklet da homeotiske gener seg?
Bilde 11. Heterozygot genpar
ND fastholder at slike gener utviklet seg ved å kode primitive tilpasninger som gjenstår å bli oppdaget, men dette er ad-hoc spekulasjoner. Bunnlinjen er at hvert nytt bevis demonstrerer universaliteten til homeotiske gener, og således deres uavhengighet av enhver tilpasning, noe som gjør at deres nærvær i en påstått felles stamfar mer vanskelig for ND å forklare. De samme oppdagelser som gjorde evo-devo så spennende, legger seg nå til lista med vanskeligheter for teorien deres.
Men hva om homeotiske gener er produktet av ID? I det tilfellet er det å finne de samme homeotiske gener tvers over ulike dyre-rekker og klasser, har sin parallell i det en finner i ordinære ingeniør-praksiser: heller enn å gjenoppfinne hjulet, gjenbruker ingeniører eksisterende design. Akkurat som en ingeniør ikke ville bli overrasket over å finne lignende tenningsbrytere i ulike motor-typer, produsert av samme forhandler, så trenger ikke biologer som innrømmer design bli overrasket over å finne lignende homeotiske gener i ulike typer av dyr. Gitt en design hypotese, så utgjør det god logikk hvorfor noen sekvenser inkorporeres i en vid variasjon av ulike dyre-rekker/klasser, som ellers viser få likheter. Til tross for manglende detaljer i både ID og ND for akkurat hvordan gener ble designet eller utviklet seg, så er ID mer konsistent enn ND med nylige oppdagelser i utviklingsgenetikk.
Referanser:
Bilde 12. Funksjonell protein-form
 10. E.J. Ambrose, The Nature and Origin of the Biological World (New York: Wiley Halated, 1982), 26
10. E.J. Ambrose, The Nature and Origin of the Biological World (New York: Wiley Halated, 1982), 26
11. S.C. Morris, Life's Solution: Inevitable Humans in a Lonely Universe (Cambridge: Cambridge University Press, 2003), 19-20
12. Ulike tripletter kan kode for samme aminosyre
13. Th. Dobxhansky, Genetids and the Origin of Species, New York: Columbia University Press, 1951), 59
14. Protein science 13 (2004): 1-14
15. Darwin, Origin of Species, 6.utg. kap. 7 http://www.literature.org/authors/darwin-charles/the-origin-of-species-6th-edition/chapter-07.html
16. Det hender også at tilpasnings-pakkene tar form av ikke-reduserbar kompleksitet i form av molekylære maskiner
18. Norbert Wiener, Cybernetics: or Control and Communication in the Animal and the Machine, 2.utg (Cambridge, MAss. MIT Press, 1961), 132
19. Ambrose, Nature and Origin og the Biological World, 120
20. E.B. Ford, Ecological Genetics (London: Chapman and Hille, 1971
21. Ambrose, Nature and Origin og the Biological World, 123
22. Ambrose, Nature and Origin og the Biological World, 143
24. Science 298 (1.Nov. 2002): 953
25. Science 298 (1.Nov. 2002): 953
26. W. Bateson, Materials for the Study of Variation Treated with Especial Regard to Disconinuity in the Origin of Species (London: MacMillan, 1894)
27. Fra et prinsipp i vitenskapelig resonnement av J. St. Mill: For å forklare en forskjell i virkning, må en identifisere en forskjell i årsak.
Oversettelse og bilder ved Asbjørn E. Lund