Ulikheter i gen-funksjoner
 (Utvalgt og oversatt fra 'More than a Monkey', Jeffrey Tomkins, PhD; Kap. 9)
(Utvalgt og oversatt fra 'More than a Monkey', Jeffrey Tomkins, PhD; Kap. 9)
I tillegg til hoved-ulikhetene som finnes mellom menneske og sjimpanser med hensyn til kromosom-antall, orfan-gener, ufullstendige slektslinjer, og ulikheter på genom-nivå, så er det også forskjeller i hvordan gener er utnyttet eller uttrykt.
Nye teknologier har satt vitenskapsmenn i stand til å sammenligne tusenvis av gener, mellom ulike typer av organismer, for å se om de slås av/på på lignende måte, hvor mye av gen-produktet blir produsert i form av RNA (uttrykks nivå), og hvilke gener de er assosiert med i form av nettverk. Selvsagt er disse data avhengig av type av organ (vev), alder på vevet, og ulike stimuli som påvirker organismen og dens vev. Vitenskapsfolk finner mange ulike, signifikante forskjeller i form av gen-uttrykk mellom mennesker og sjimpanser, og i mange tilfeller har ikke disse noen klar evolusjonær forklaring, og motsetter seg det darwinistiske dogmet.
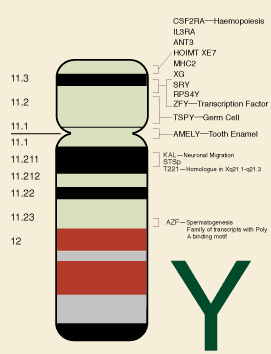
Bilde 1. Y-kromosom hos mennesket
Protein ulikheter og likheter
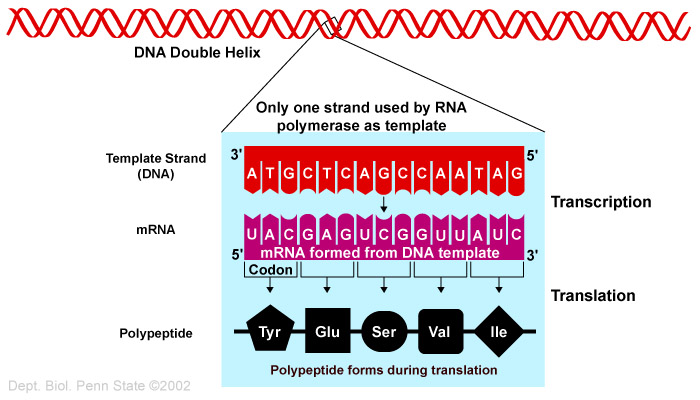 Sluttproduktet til det vi tradisjonelt refererer til som et gen, er et protein, som uttrykkes i form av kjeder av spesielt ordnede aminosyrer, som koder for dette genet. Etter at proteiner er oversatt fra m-RNA (messenger RNAs), som kopieres og skrives av fra gener, så foldes de til spesifikke, forutbestemte, 3-dimensjonale former. Proteiner kan også kombineres med andre proteiner, metall-ioner, karbohydrater og RNA til å danne større komplekse 'maskiner'. Men ikke alle RNA-produkt fra genomet fører til proteiner. I stedet benyttes de til både funksjonelle og strukturelle formål, og havner i en forvirrende gruppe av kategorier. Faktisk er det over dobbelt så mange av lange, ikke-kodende RNA gener, som det er av protein-kodende gener i det menneskelige genom. Mange av disse er endog mer celle -og vevs-spesifikke i sitt uttrykk enn protein-kodende gener.
Sluttproduktet til det vi tradisjonelt refererer til som et gen, er et protein, som uttrykkes i form av kjeder av spesielt ordnede aminosyrer, som koder for dette genet. Etter at proteiner er oversatt fra m-RNA (messenger RNAs), som kopieres og skrives av fra gener, så foldes de til spesifikke, forutbestemte, 3-dimensjonale former. Proteiner kan også kombineres med andre proteiner, metall-ioner, karbohydrater og RNA til å danne større komplekse 'maskiner'. Men ikke alle RNA-produkt fra genomet fører til proteiner. I stedet benyttes de til både funksjonelle og strukturelle formål, og havner i en forvirrende gruppe av kategorier. Faktisk er det over dobbelt så mange av lange, ikke-kodende RNA gener, som det er av protein-kodende gener i det menneskelige genom. Mange av disse er endog mer celle -og vevs-spesifikke i sitt uttrykk enn protein-kodende gener.
Bilde 2. DNA og RNAs funksjoner i cellen
Det er estimert at mindre enn 5% av det menneskelige genom, består av protein-kodende DNA-sekvenser (eksoner). Ikke desto mindre er det her vitenskapsmenn har fokusert, på protein-kodende sekvenser -som er er mest mulig like, i sin iver etter å finne DNA som er likt mellom sjimpanse og menneske. Selv i denne smale strategien, er det dukket opp resultater som viser at menneske og sjimpanse klart er ulike. Fokuset har vært på noe som kalles 'ortologe' (lignende) proteiner, og antas å ha evolvert fra ett enkelt opphavs-gen. En av de mest siterte studiene av dette slaget, ble publisert omtrent samtidig som sjimpanse gen-sekvensen. Denne studien sammenlignet 127 ortologe proteiner fra sjimpanse og menneske. Av disse var det bare 20% (25 proteiner) som viste identisk aminosyre-sekvens, og 80% var ulike -med ganske små endringer i de fleste tilfelle. Problemet var at et utvalg på 127 proteiner er veldig lite, av de over 1 million som mennesket innehar, og primært var de kun fysiologiske proteiner som finnes i de fleste pattedyr. Det er likevel viktig å være klar over at små forskjeller mellom proteiner, kan produsere relativt store variasjoner i deres tre-dimensjonale form og funksjon.
 En av de største protein-sammenlignende studiene, kom i 2006, der forskere identifiserte 1.480 menneskelige protein-kodende gener, som ikke hadde noen lignende (ortologe) gener i sjimpanse. Det er et signifikant antall gener, når forskere nå tror det er godt over 27.000 protein-kodende gener i det menneskelige genom. Det totale antall proteiner som nå er oppdaget i mennesket, utgjør nå over en million, grunnet alternativ spleising av kodende segmenter, som kalles eksoner.
En av de største protein-sammenlignende studiene, kom i 2006, der forskere identifiserte 1.480 menneskelige protein-kodende gener, som ikke hadde noen lignende (ortologe) gener i sjimpanse. Det er et signifikant antall gener, når forskere nå tror det er godt over 27.000 protein-kodende gener i det menneskelige genom. Det totale antall proteiner som nå er oppdaget i mennesket, utgjør nå over en million, grunnet alternativ spleising av kodende segmenter, som kalles eksoner.
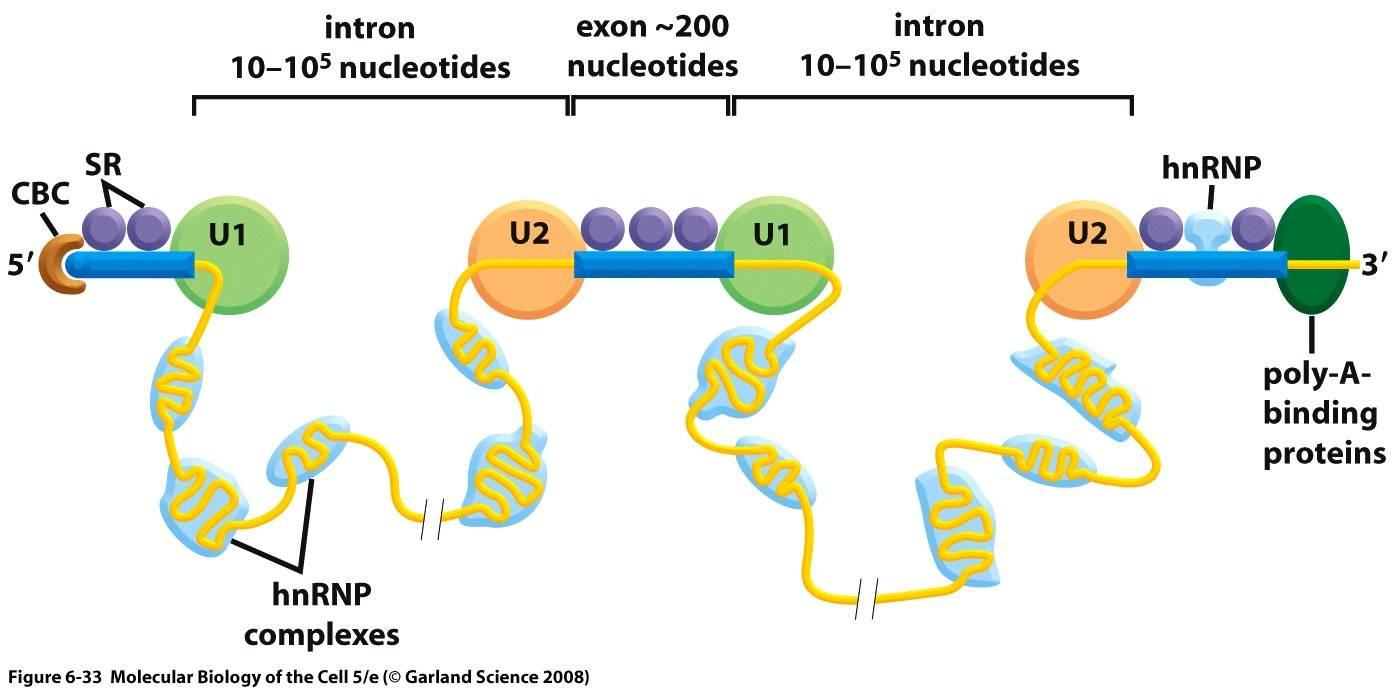
Bilde 3. naturlig spleising av gener
Forskjeller i gen-uttrykk
Proteiner produseres fra m-RNA avskrifter, kopiert fra gener. Før de brukes til å produsere et protein, så prosesseres avskriften, noe som involverer spleising av protein-kodende (ekson) segmenter i RNA. Spleising er prosessen der de ikke-kodende mellomliggende segmenter (introner) fjernes, og kodende (ekson) regioner i genet lenkes sammen for å generere moden RNA, som er oversatt til proteiner. Multippel spleising konfigurasjon av avskrifter for ett enkelt gen, kan generere mange proteinvarianter, avhengig av genet. Varianter av spleising kommer inn under meget komplekse genetiske kontroll-mekanismer. Faktisk har forskning vist at alternativ spleising er betydelig ulik mellom mennesker og sjimpanser, selv for tilsynelatende like gener (3). Mens et betydelig antall av gener mellom mennesker og sjimpanser deler likhet (orthologous genes), så synes faktorene som kontrollerer hvordan de uttrykkes, å spille en mye større rolle i de observerte ulikhetene (4,5). Selv om ulike typer av organismer deler de samme gentypene, så er det ulikt hvordan genene benyttes. Vi vet nå at proteinkodende gener bare er basis-enhetene til genomet i en 'innsjø' av metadata som styrer bruken deres.
 En studie fra 2010 bekreftet de store forskjellene i uttrykk sammen med linken mellom hjerne og stoffskifte-gener i et større datasett (7). Til støtte for denne forbindelsen mellom forbundne genetiske data, så påviste en publisert studie fra 2011 nivåmålinger i celle-metabolismer mellom menneske og sjimpanse-hjerner. Den fant hovedforskjeller mellom 77% av nivåene for hvert av disse molekylene (9). Om mennesker skulle utviklet seg fra aper, så ville ikke bare hjernegener ha måttet endret seg merkbart, men også gener fra multiple nettverk og andre relatert til stoffskifte, for å støtte de multippelt-avanserte funksjonene til hjernen. Menneskelig hjerne trenger enorme mengder med tilført celle-energi, og koordinert stoffskifte-støtte fra resten av kroppen. Selvsagt måtte disse endringene skje i perfekt utført tandem, som er en høyst kompleks og umulig evolusjonær bedrift.
En studie fra 2010 bekreftet de store forskjellene i uttrykk sammen med linken mellom hjerne og stoffskifte-gener i et større datasett (7). Til støtte for denne forbindelsen mellom forbundne genetiske data, så påviste en publisert studie fra 2011 nivåmålinger i celle-metabolismer mellom menneske og sjimpanse-hjerner. Den fant hovedforskjeller mellom 77% av nivåene for hvert av disse molekylene (9). Om mennesker skulle utviklet seg fra aper, så ville ikke bare hjernegener ha måttet endret seg merkbart, men også gener fra multiple nettverk og andre relatert til stoffskifte, for å støtte de multippelt-avanserte funksjonene til hjernen. Menneskelig hjerne trenger enorme mengder med tilført celle-energi, og koordinert stoffskifte-støtte fra resten av kroppen. Selvsagt måtte disse endringene skje i perfekt utført tandem, som er en høyst kompleks og umulig evolusjonær bedrift.
Bilde 4. Fra DNA til ferdig dyr
I 2005 undersøkte en annen gruppe ulikheter i gen-uttrykk for en rekke organer mellom menneske og sjimpanse (10). I overensstemmelse med andre studier, var forskjeller for hvordan gener ble uttrykt, høyst signifikante for hjernen og andre organer som nyrer og lever, samt for testiklene. Dette ble senere bekreftet av den store ulikheten i Y-kromosom mellom menneske og sjimpanse. En annen metode for å identifisere gen-uttrykking, er å identifisere overlappende sett og del-sett blant moduler av gener. Gener blir ofte uttrykt i moduler eller grupper, relatert til ulike celle-funksjoner. Det overlappende genmengden til interrelaterte moduler, kalles nettverk-forbindelser. En studie som nyttiggjorde dette begrepet for å teste likhet mellom menneske og sjimpanser, fant at 17,4% av nettverksforbindelsene for gener, var spesifikke for mennesket og ble ikke funnet i sjimpanser (4).
Regulerende DNA-ulikheter
Studier om regulerende sekvenser, handler om hvordan DNA-egenskaper ved genomet kontrollerer hvordan gener uttrykkes. En slik studie sammenligner aktivator-regioner i homologe gener i sjimpanse, makaker og  menneske. Aktivatorer er seksjoner av DNA som regulerer genet og spiller en stor rolle i regulering av dets uttrykks-nivå, ved å binde til proteiner, og kalles transkripsjonsfaktorer. Av de undersøkte homologe genene, ble 575 menneskelig gener funnet å være meget ulike fra de i sjimpanser (11). Mange av disse DNA-ulikhetene var relatert til gener som kontrollerer nervecellers utvikling. Men overraskende nok, var andre relatert til mer basis stoffskifte-aktiviteter, f.eks. i forbindelse med karbohydrat. Genregulering i forbindelse med proteiner som binder DNA og koder avskrivnings (transcription) faktorer, er meget sentrale i dannelsen av en organisme. Flere studier har forsøkt å evaluere likheten i dette mellom sjimpanser og mennesker. I en nylig undersøkelse om emnet, erklærte forfatterne at: "Funksjonelle analyser av genom-skanning har avslørt akselererende sekvens-evolusjon av transkripsjons-faktorer, spesielt 'zinc finger' proteiner. Transkripsjonsfaktorer som inneholdt denne, ble også funnet å være overrepresentert blant gener som nylig har oppstått i den menneskelige linjen gjennom dupliseringer." (5)
menneske. Aktivatorer er seksjoner av DNA som regulerer genet og spiller en stor rolle i regulering av dets uttrykks-nivå, ved å binde til proteiner, og kalles transkripsjonsfaktorer. Av de undersøkte homologe genene, ble 575 menneskelig gener funnet å være meget ulike fra de i sjimpanser (11). Mange av disse DNA-ulikhetene var relatert til gener som kontrollerer nervecellers utvikling. Men overraskende nok, var andre relatert til mer basis stoffskifte-aktiviteter, f.eks. i forbindelse med karbohydrat. Genregulering i forbindelse med proteiner som binder DNA og koder avskrivnings (transcription) faktorer, er meget sentrale i dannelsen av en organisme. Flere studier har forsøkt å evaluere likheten i dette mellom sjimpanser og mennesker. I en nylig undersøkelse om emnet, erklærte forfatterne at: "Funksjonelle analyser av genom-skanning har avslørt akselererende sekvens-evolusjon av transkripsjons-faktorer, spesielt 'zinc finger' proteiner. Transkripsjonsfaktorer som inneholdt denne, ble også funnet å være overrepresentert blant gener som nylig har oppstått i den menneskelige linjen gjennom dupliseringer." (5)
Bilde 5. Kromosomlikhet menneske-sjimpanse
Typen av transkripsjonsfaktorer kalt: 'Zinc finger' er den største undergruppen av slike proteiner. Da forfatterne påsto at disse "nylig hadde opstått i den menneskelige linjer," betyr det at de hadde oppstått plutselig, og ikke hadde noen evolusjonær historie. Og når de erklærer at de var duplisert, innebærer det bare at deler av noen gener i genomet, deler seksjoner med liknende kode. Ut fra evolusjonære forutsetninger fortolkes det som at genene med lignende elementer ble duplisert fra andre pre-eksistente gener. Hva mange biologer ikke tar i betraktning, er at visse seksjoner av kode blir gjenbrukt. Det skjer i lange data-programmer, og indikerer design og konstruksjon. I et dataprogram kan en ikke bare 'klippe og lime' programkode på måfå, og forvente at det kommer noe virksomt ut av det. Genomet virker på samme måte, og tilfeldig innsetting av fremmede gener i plante og dyr-genomer har, gjennom genteknologi, demonstrert at dette er tilfelle.
 Et annet område av funksjonelle ulikheter mellom menneskets og sjimpanses gen-uttrykk, har å gjøre med ikke-kodende DNA som uttrykkes utenfor protein-kodende regioner. Det er et meget komplisert emne, og involverer mange typer av både korte og lange ikke-kodende RNA. Mange av dem virker i kjernen, mens andre eksporteres til cellens cytoplasma. En type av kort RNA, som regulerer mRNA etter at de 'skrives-av', kalles mikro-RNA. Det er funnet mange ulikheter i uttrykksnivåer for mikroRNA, mellom menneske og sjimpanse, inkludert mikroRNA som kun er funnet i menneske. (5)
Et annet område av funksjonelle ulikheter mellom menneskets og sjimpanses gen-uttrykk, har å gjøre med ikke-kodende DNA som uttrykkes utenfor protein-kodende regioner. Det er et meget komplisert emne, og involverer mange typer av både korte og lange ikke-kodende RNA. Mange av dem virker i kjernen, mens andre eksporteres til cellens cytoplasma. En type av kort RNA, som regulerer mRNA etter at de 'skrives-av', kalles mikro-RNA. Det er funnet mange ulikheter i uttrykksnivåer for mikroRNA, mellom menneske og sjimpanse, inkludert mikroRNA som kun er funnet i menneske. (5)
Bilde 6. Introner-Ekson-bindings protein
I en oversikt over studier som sammenlignet menneske og ape -like RNAs, så erklærte forfatterne at det er identifisert et stort antall av nye ncRNA-avskriftskoder, hvorav mange av dem hadde menneske-spesifikke uttrykk (5). I en nylig studie av mennesker, sjimpanser og makaker, evaluerte forskerne 1.898 ulike linkRNA-gener i ni ulik vevstyper. 80% av mennesklig linkRNA hadde tilsvarende motparter uttrykt i sjimpanse, og bare 63% i makaker(14). Som i mange andre studier av underklasser av lncRNA, så er disse typene betydelig mer arts- (taksonomisk) begrenset enn proteinkodende eksoner, og viser seg å være nøkkelegenskaper i genomet til å markere genetiske grenser mellom hva kreasjonister ville kalle 'skapte slag'.
Ulikheter i genenes nabolag
Et signifikant antall gener som opererer innen det samme fysiologiske nettverk i cellen, lokaliseres også tett sammen i genomet, og har delte uttrykksformer mellom ulike typer av organismer. Men skyldes dette evolusjonær progresjon i genomet, eller gjentatte design-mønstre iboende i konstruerte systemer? En nylig studie sammenlignet geners nabolag mellom ulike grupper som mennesker, sjimpanser, mus, rotter, kylling, sebrafisk og bananfluer (15). Det var større ulikhet mellom ulike dyregrupper i sammenlignbare gen-naboområder, enn for genomet generelt. Det gjaldt særlig gen-rekkefølge mellom to ulike organismer, og tar også i betraktning nærvær/fravær av gener når to ulike organismer blir sammenliknet. Forfatteren av dette studiet erklærte: 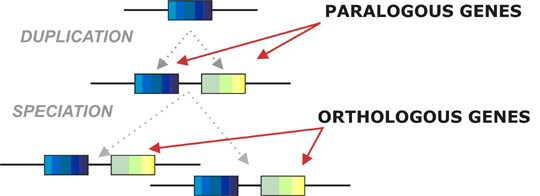 "Overraskende nok er genene som finnes i funksjonelle naboområder og delt av ulike organismer, ikke nødvendigvis lik hverandre (orthologous). Når to arter deler funksjonelle naboområder, så kan genene som former disse, være ulike i hver art (15).
"Overraskende nok er genene som finnes i funksjonelle naboområder og delt av ulike organismer, ikke nødvendigvis lik hverandre (orthologous). Når to arter deler funksjonelle naboområder, så kan genene som former disse, være ulike i hver art (15).
Bilde 7. 'Ortologe' gener
En annen oppdaget hovedforskjell mellom artene kom fra sammenligning av klynger av gener på kromosomer (gene neigborhoods), som tilsynelatende hadde like funksjoner i cellen (16). Den store overraskelsen var at det samme funksjonelle området i et signifikant antall tilfelle, var sammensatt av ulike gener. For 18% av genene som var like mellom mennesker og sjimpanser, ble det funnet at det eksisterte store forskjeller i gen-landskapet omkring dem, som de kalte 'endrede gen-landskap'. Til tross for store likheter i homologi mellom de angjeldende gener hos sjimpanse og menneske, så var det genlandskapet omkring dem ulikt, eller artsspesifikt. Disse ulikheter i genlandskap, ble også funnet å være assosiert med ulikheter av hvordan genene ble uttrykt mellom meneske og sjimpanse. Det er altså ikke nok kun å se på homologe gener. Den genomiske konteksten som de lokaliseres i, og hvilken funksjon de innehar, er også viktig å betrakte, noe som ikke støtter makro-evolusjon. I stedet ser vi gjentatt bruk og variasjon av visse design-konsept, i mosaikk-mønstre mellom skapninger, der felles avstamning ikke brukes som forklaring.
Konklusjon: Gen-uttrykk motsier menneskelig evolusjon
Mange lignende gener mellom menneske og sjimpanse uttrykkes meget ulikt, og uttrykksprofilen bærer ikke spor av seleksjon eller å ha blitt utviklet. Gener knyttet til transport, hjerne, celle-signalering, testikler og mange stoffskifte-funksjoner viser distinkt ulike uttrykksformer. Disse gener forekommer i utstrakte nettverk, og må være presist koordinert både i disse og mellom organer over hele kroppen. Mange uttrykte gener i mennesket, er ikke funnet i sjimpanse. Forskning har klart vist et mønster av ikke-reduserbar kompleksitet i genomet, som opptrer plutselig og fullt integrert i mennesker ulikt fra sjimpanser.
Utvalg av stoff og bilder ved Asbjørn E. Lund