Darwins evolusjonstre
 (Utdrag fra boka 'Icons of Evolution' av J. Wells)
(Utdrag fra boka 'Icons of Evolution' av J. Wells)
Da Darwin skrev 'Artenes Opprinnelse' i 1859 var Kambrium den eldste geologiske periode en hadde funnet fossiler fra. Darwin trodde at alle organiske vesener som har levd på jorden, kunne nedstamme fra en og samme ur-organisme. I 'Artenes Opprinnelse' var det kun én illustrasjon, og det er en tegning som viser hvordan livet er forgrenet som resultat av en prosess han kaller nedarving med tilpasning. Darwin forestilte seg at livets historie kunne symboliseres ved et tre med en altomfattende felles stamfar ved dets rot og nåtidens mange ulike arter som det spirende kvister. Denne tegningen kalte han
Av alle evolusjonære ikoner er livets 'stamtre' det mest betydningsfulle, fordi nedstamning fra en felles forfader er selve fundamentet for Darwins teori. Likevel var Darwin klar over og forskerne har nylig bekreftet, at det eldste fossilmateriale vender opp ned på livets evolusjonære stamtre. For 10 år siden håpet evolusjonister at det molekylære forskningsmaterialet ville kunne redde treet, men nyere oppdagelser har fått det håpet til å rakne. Selv om ikke biologibøkene sier det, er Darwins evolusjonstre revet opp med roten. 'Livets mektige tre' (opprinnelig tegning finnes ikke i høyere oppløsning).
Darwins evolusjonstre
Hvis alle livsformer stammer fra en felles forfader, hvorfor utviser de så stor diversitet? Dyre og planteforedlere kan endre på eksisterende former ved å utvelge enkelte egenskaper til 'videreforedling' eller masseformering. Darwin hevdet at en tilsvarende prosess fant sted i naturen. Hvis en del av en naturlig populasjon blir utsatt for én type livsbetingelser, og andre deler blir utsatt for noen andre, ville 'den naturlige seleksjon' kunne endre ulike undergrupper i hver sin retning. Om det var tid nok, ville en enkelt art kunne danne atskillig nye varianter. Darwin trodde at hvis disse nye varianter fortsatte å splitte seg opp, ville de etter hvert danne nye arter. I stamtre-illustrasjonen er tiden loddrett akse, med 1000 generasjoner mellom hver vannrette strek. Den vannrette aksen viser forskjeller mellom organismene. I følge Darwin skulle bittesmå variasjoner i starten bli forsterket over generasjonenes løp og bli så store at de ville atskille den ene art fra den andre. F.eks. skal ur-organisme: 'I' på livstreet etter 14.000 generasjoner ha utviklet seg til 6 arter. Den lange tiden er som kjent nødvendig, men er den tilstrekkelig?
Det biologiske klassifikasjonssystem ble utviklet av Carl von Linné et århundre før Darwin, og det brukes stadig av de fleste biologer. Her er organismene hierarkisk fordelt i grupper, alt etter likheter og forskjeller. Viser et eks ut fra Fig 3-2, siden vi trenger klassifikasjons-begreper til dokumentasjon i det videre.  I følge Darwins teori har menneske og bananflue hatt samme stamfar.
I følge Darwins teori har menneske og bananflue hatt samme stamfar.
'Livstreet' er ment å vise utviklingen fra individ til arter, fra arter til slekter osv opp fra klasse-til rekke nivå. Men det er ikke plass på 'Livstreet' til at ulikheter på rekkenivå kan være til stede fra første begynnelse. Men det er nettopp hva fossilmaterialet viser:
Darwin og fossilmaterialet
Da Darwin skrev 'Artenes Opprinnelse', var de eldste fossiler en kjente til fra den geologiske periode en kaller Kambrium. Perioden har fått navn etter klipper i Cambria i Wales (fig3-3). Men Kambriums sammensetning av fossiler passer ikke med Darwins teori. Kambrium begynner med en rekke fullt utviklede dyre-klasser og -rekker. Det biologiske hierarkis høyeste nivå er med andre ord til stede helt fra begynnelsen. Darwin var klar over dette, og anså det være en vesentlig vanskelighet for sin teori. Han forutsatte imidlertid at .."før de nederste lag i Kambrium var avsatt, var det lange perioder forløpt ..[i hvilke] verden har vrimlet med forskjellige levende vesener."
Han formodet at bergarter eldre enn Kambrium var i den grad forvandlet av trykk og temperatur at alle spor etter fossiler er blitt ødelagt. Og derfor 'ser det ut som' de store dyregruppene plutselig framtrer i Kambrium. Han forutsatte at den fortsatte jakt på fossiler i det minste ville kunne levere noe av det manglende forskningsmateriale. Siden da har ytterligere forskning avslørt mange fossilholdige lag eldre enn Kambrium. Det har ført til at vår forståelse av hva som skjedde før Kambrium er vesentlig bedre enn Darwins. Paleontologer har videre funnet bergarter fra Kambrium i Canada, Grønland og Kina, vedsiden av dybhavsboringer etter olje til havs..
Det rare er at denne voldsomt økende viten om kambriske og pre-kambriske fossiler snarere har forstørret enn forminsket Darwins problem. Mange paleontologer er i dag overbevist om at store dyregrupper faktisk har oppstått ganske plutselig i tidlig Kambrium. Fossilmaterialet står så sterkt og begivenheten er så dramatisk, at den i dag kalles den kambriske eksplosjon, eller 'biologiens big bang'.
Den kambriske eksplosjon
 I Afrika og Australia har geologer rapportert om sedimentbergarter som er over tre milliarder år gamle og inneholder fossiler fra éncellede organismer. En har funnet litt yngre sedimentbergarter, som inneholder fossile 'stromatolitter', lagdelte puteformede strukturer av fotosyntetiske bakteriesamfunn. Det er funnet spesielle flercellede organismer tidligere enn Kambrium, først funnet i Ediacara Hills i Syd-Australia, men senere flere steder. Noen paleontologer hevder at Ediacara-fossilene er forfedre til dyr som senere oppsto i Kambrium. Andre hevder at de er så ulike at de bør plasseres i sitt eget rike.
I Afrika og Australia har geologer rapportert om sedimentbergarter som er over tre milliarder år gamle og inneholder fossiler fra éncellede organismer. En har funnet litt yngre sedimentbergarter, som inneholder fossile 'stromatolitter', lagdelte puteformede strukturer av fotosyntetiske bakteriesamfunn. Det er funnet spesielle flercellede organismer tidligere enn Kambrium, først funnet i Ediacara Hills i Syd-Australia, men senere flere steder. Noen paleontologer hevder at Ediacara-fossilene er forfedre til dyr som senere oppsto i Kambrium. Andre hevder at de er så ulike at de bør plasseres i sitt eget rike.
Like før Kambrium er det også funnet to tegn til på forekomster av flercellede dyr: Det ene er Small Shelley Fauna (SSF), oversatt småskalldyrs-fauna. SSF er små, forsynt med skall og har de første forekomster av skjelettliknende deler, men ikke lik noen nålevende gruppe. Det andre er sporfossiler (gang og avtrykk) som tilsynelatende er laget av en flercellet orm. Men bortsett fra sistnevnte finnes det ikke fossilmateriale som forbinder dyr fra Kambrium med organismer som går forut for dem. Den nå veldokumenterte prekambriske fossilhistorie stiller ikke med noe som kan ligne på Darwins lange historie om gradvis oppsplitting.
I løpet av 1980-årene skjedde det noe som gjorde at noen forskere fikk øynene opp for betydningen av dette. Da ble fossiler fra det tidligere funne Burgess Shale i Canada anlysert på ny. I tillegg ble det oppdaget to andre lokaliteter med fossiler i stil med Burgess Shale: Sirius passet på Nord-Grønland og Chengjiang i Sør-Kina. Fossilene fra Chengjiang syntes å være de eldste, fra Kambrium de også, og best bevarte. De omfatter hva som kan være de første virveldyr. Disse funn er blitt gjort av en anerkjent kinesisk 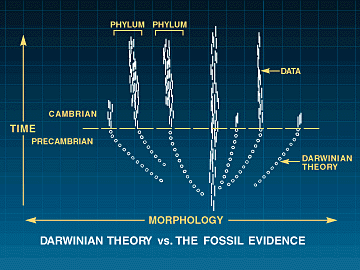 paleontolog ( paleontolog: en som forsker på bergarter, dyre- og plante-fossiler). Da han besøkte USA for å forelese om funnene, påpekte han at 'opp-ned' mønsteret fra Kambrium motsier darwinismen. Dette ble ignorert av forskerne der, og kommentert av gjesten slik: 'I Kina må en gjerne kritisere Darwin, men ikke regjeringen. I USA må man gjerne kritisere regjeringen, men ikke Darwin.'
paleontolog ( paleontolog: en som forsker på bergarter, dyre- og plante-fossiler). Da han besøkte USA for å forelese om funnene, påpekte han at 'opp-ned' mønsteret fra Kambrium motsier darwinismen. Dette ble ignorert av forskerne der, og kommentert av gjesten slik: 'I Kina må en gjerne kritisere Darwin, men ikke regjeringen. I USA må man gjerne kritisere regjeringen, men ikke Darwin.'
Perioden Kambrium ligger antagelig ca 550 millioner år tilbake i tid. Den betydelige forøkelsen av fossiler fant sted for ca 530 millioner år siden og varte fra 5 til høyst 10 millioner år. Det tilsvarer 1-2% av jordas levealder. Den kambriske eksplosjon gav opphav til de fleste av de dyrerekker som eksisterer i dag, foruten en del som er utdødde. I følge paleontologene James Valentine, Steanley Awramik, Phillip Signor og Peter Sadler, "er den fossile lagseries helt klart mest spektakulære fenomen, den plutselige oppdukking av oppsplitting i mange nålevende og utdødde rekker" ved Kambriums begynnelse. Mange dyrs legemsstrukturer, (som har gjort at de kan rubriseres i ulike rekker og klasser) "har først utviklet seg på dette tidspunkt, i en periode på høyst noen få millioner år". Valentine og kolleger nådde fram til den konklusjon at "den kambriske eksplosjon har vært enda mer abrupt og omfattende enn tidligere antatt".
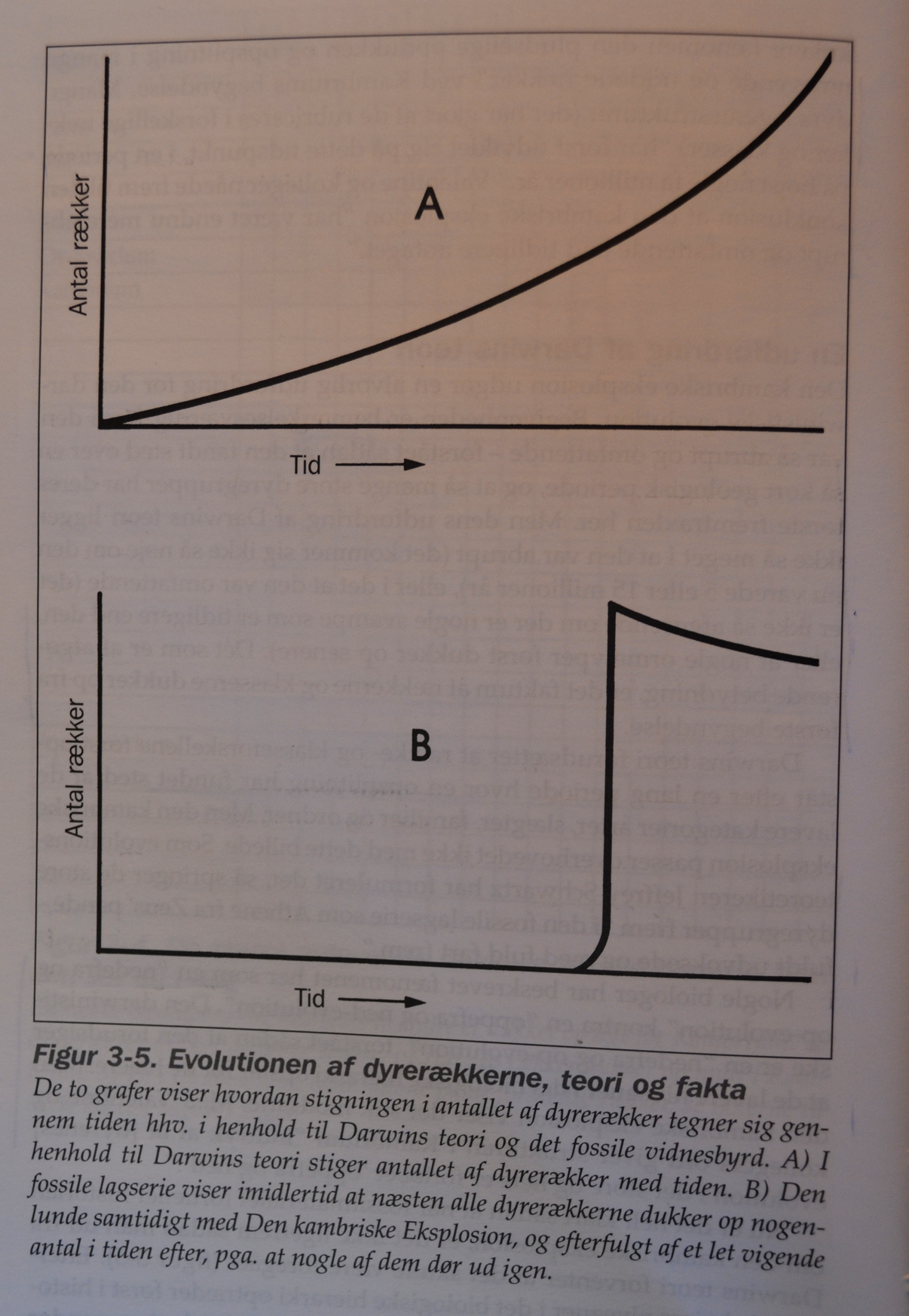 En utfordring til Darwins teori
En utfordring til Darwins teori
Det at dyre-rekkene og -klassene dukket opp så fort, i løpet av 5-10 milioner år, fullt ferdig utviklet -uten spor til den lange utviklingsperioden Darwin trengte, er en alvorlig utfordring for hans teori. Med Valentine og kollegers ord gir utviklingen i Kambrium i det store og hele å ha forløpt 'ovenfra og ned'. Ettersom de høyeste nivåene i det biologiske hierarki opptrer først i historien, kan vi si at den kambriske eksplosjon vender Darwins evolusjonstre på hodet. Ikke dessto mindre har evolusjonsbiologer villet overse dette faktum og sett etter måter å redde Darwins teori på.
Hvordan redde Darwins teori?
Med henblikk på den kambriske eksplosjon er det tre måter å redde Darwins teori på:
1) Å argumentere (som også Darwin gjorde) at tilsynelatende mangel på forfedre før Kambrium skyldes mangel på fossiler. 2) Å hevde at selv om utviklingsrekker fortsetter ned i Prekambrium, kan en ikke se det fordi stamformene ikke har dannet fossiler. Enten har de vært for små eller de har bare bestått av bløtdeler. 3) Å overtrumfe det fossile vitnesbyrd med et molekylært, hvor en ved å sammenligne nålevende former med hverandre får en indikasjon på en hypotetisk felles stamform, antagelig hundrevis av millioner år før Kambrium.
 Ad 1) En har i dag funnet tilstrekkelig gode sediment-bergarter fra sent PreKambrium og Kambrium til å overbevise paleontologene om at hvis det hadde fantes noen forfedre -og de hadde dannet fossiler, så ville de vært oppdaget nå. I følge James Valentine og Douglas Erwin er det slik at 'de serier av bergarter fra Kambrium vi har, er like så komplette og oppbygget under samme sedimentære forhold som lagserier som ligger er tettere på nåtiden. Allikevel er det slik at forfedre er "ukjente eller ubekreftede" for alle rekker eller klasser som dukket opp i den kambriske eksplosjon. "Den kambriske eksplosjon er ..for omfattende til at den kan bortforklares med defekter i den fossile lagserie." Flere ulike undersøkelser av den fossile lagserie fra Kambrium og opp til nå har bekreftet dette synet. I februar år 2000 når de britiske geologer M.J.Benton, M.A.Wills og R.Hitchin fram til flg.
Ad 1) En har i dag funnet tilstrekkelig gode sediment-bergarter fra sent PreKambrium og Kambrium til å overbevise paleontologene om at hvis det hadde fantes noen forfedre -og de hadde dannet fossiler, så ville de vært oppdaget nå. I følge James Valentine og Douglas Erwin er det slik at 'de serier av bergarter fra Kambrium vi har, er like så komplette og oppbygget under samme sedimentære forhold som lagserier som ligger er tettere på nåtiden. Allikevel er det slik at forfedre er "ukjente eller ubekreftede" for alle rekker eller klasser som dukket opp i den kambriske eksplosjon. "Den kambriske eksplosjon er ..for omfattende til at den kan bortforklares med defekter i den fossile lagserie." Flere ulike undersøkelser av den fossile lagserie fra Kambrium og opp til nå har bekreftet dette synet. I februar år 2000 når de britiske geologer M.J.Benton, M.A.Wills og R.Hitchin fram til flg.  konklusjon: "Den fossile lagseries tidligste deler er klart ufullstendige, men en kan saktens betrakte dem som pålitelige nok til å tegne de brede linjer i livets historie.
konklusjon: "Den fossile lagseries tidligste deler er klart ufullstendige, men en kan saktens betrakte dem som pålitelige nok til å tegne de brede linjer i livets historie.
(Bilder av fossilavtrykk ved siden og over fra traileren til: Darwins Doubt)
Ad 2) Er fossilene for små eller for bløte? Problemet med denne bortforklaringen er at en har funnet mikrofossiler av bakterier i bergarter som er over 3 milliarder år gamle. De nevnte funn i Ediacara Hills besto kun av bløtdeler. Det samme gjelder funnene i Burgess Shale: mange av funnene består av bløtdeler. En kan i følge Conway Morris også se indre organer som innvoller eller muskler i disse funnene. Så manglende funn skyldes bestemt ikke at de har bestått av kun bløtdeler eller vært for små. I følge Schopf er det nå bred enighet om at det forholder seg slik.
 Ad 3) Den tredje måte evolusjonsbiologer har prøvd demontere den kambriske eksplosjon på, er ved å hevde at molekylært forskningsmateriale fra nålevende skapninger peker i retning av en felles stamfar til dyrerekkene, hundrevis av millioner år før Kambrium. For å forstå dette argumentet og hvorfor det ikke 'holder vann', er vi nødt til å se nærmere på 'molekylær fylogenese' (kommer i slutten av november 2013)
Ad 3) Den tredje måte evolusjonsbiologer har prøvd demontere den kambriske eksplosjon på, er ved å hevde at molekylært forskningsmateriale fra nålevende skapninger peker i retning av en felles stamfar til dyrerekkene, hundrevis av millioner år før Kambrium. For å forstå dette argumentet og hvorfor det ikke 'holder vann', er vi nødt til å se nærmere på 'molekylær fylogenese' (kommer i slutten av november 2013)
Molekylær fylogenese
Fylogenese: En fylogenese er en gruppe organismers evolusjonære historie. Inntil nylig utledet en fylogenesen av anatomiske og fysiologiske trekk (antall lemmer, varmblodig/kaldblodig etc). Etter at den Molekylær fylogenesen har gjort sitt inntog, baseres mange fylogeneser nå på DNA- og protein sammenligninger. Molekylær fylogenese utforsker altså slektskapsforholdene mellom arter eller grupper av arter. Ordet kommer av de greske røttene φύλον (fylon), «stamme» og γένεσις (genesis), «dannelse».
Alle levende organismer fra bakterier til mennesker inneholder DNA. Et DNA-molekyl er en lang kjede som blant annet består av ulike kombinasjoner av fire baser eller nukleotider som for korthets skyld kalles (A,T,C og G). Rekkefølgen av disse basene bestemmer  hvilke aminosyrene som inngår i en organismes mange ulike proteiner. Under replikasjonen blir rekkefølgen av baser kopiert til et nytt DNA-molekyl. Men molekylære tilfeldigheter eller mutasjoner kan noen ganger gjøre kopien ulik modermolekylet. Derfor kan organismer inneholde DNA-molekyler, og dermed også proteiner, som i noen grad avviker fra forfedrenes DNA og proteiner.
hvilke aminosyrene som inngår i en organismes mange ulike proteiner. Under replikasjonen blir rekkefølgen av baser kopiert til et nytt DNA-molekyl. Men molekylære tilfeldigheter eller mutasjoner kan noen ganger gjøre kopien ulik modermolekylet. Derfor kan organismer inneholde DNA-molekyler, og dermed også proteiner, som i noen grad avviker fra forfedrenes DNA og proteiner.
I 1962 hevdet biologene Emile Zuckerkandl og Linus Pauling at sammenligninger av DNA-rekkefølger og deres protein-produkter kan brukes til å avgjøre hvor tett beslektet organismer er. De organismer hvis proteiner avviker bare med noen få baser, er sannsynligvis nærmere beslektet i evolusjonær forstand, enn dem som har flere avvik. Hvis mutasjonene er akkumulert i samme takt over tid, kan antall ulikheter mellom organismene brukes som et molekylært ur, som indikerer hvor lenge det er siden deres DNA skulle ha vært identisk. (Fig.3-6)
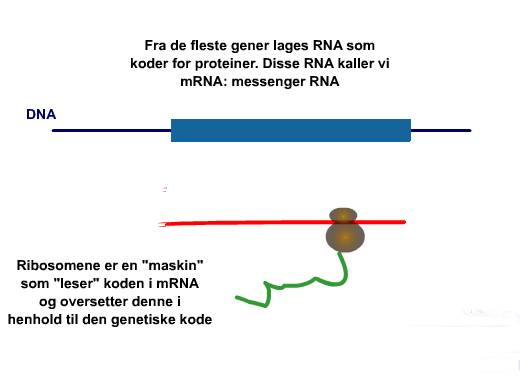 Da det ble raskere teknikker for analyse av DNA-sekvenser, ble det mer alminnelig å analysere genkodene enn proteinene, slik det tidligere var gjort. Utover DNA og proteiner, inneholder også alle organismer RNA, som er tett kjemisk beslektet med DNA. RNA bistår i å overføre informasjon fra DNA til proteinsekvensene. En del av denne prosessen beror på bittesmå organeller i cellen som kalles ribosomer. Se animasjon av oppbygningen her. (Bilde: Ribosom)
Da det ble raskere teknikker for analyse av DNA-sekvenser, ble det mer alminnelig å analysere genkodene enn proteinene, slik det tidligere var gjort. Utover DNA og proteiner, inneholder også alle organismer RNA, som er tett kjemisk beslektet med DNA. RNA bistår i å overføre informasjon fra DNA til proteinsekvensene. En del av denne prosessen beror på bittesmå organeller i cellen som kalles ribosomer. Se animasjon av oppbygningen her. (Bilde: Ribosom)
Ribosomene inneholder ribosom-RNA, som forkortes rRNA. Siden 1980 har de DNA-sekvenser som koder for rRNA tilveiebragt mye av informasjonen som er samlet om molekylær fylogenese. Om teorien for å sammenlikne DNA-sekvenser er enkel, er det vanskeligere i praksis. Et gitt DNA-segment kan inneholde tusenvis av baser (nukleotider). Bare det å få dem stilt opp i rekkefølge for sammenligning kan være krevende. Og ulike oppstillinger kan gi ulike resultater. Ikke desto mindre er det trukket konklusjoner ut fra molekylær fylogenese som har betydning for utvikling av arter. Særlig har det interesse i forbindelse med Kambrium perioden, som vi tidligere har sett på.
 Molekylær fylogenese og den kambriske eksplosjon
Molekylær fylogenese og den kambriske eksplosjon
Oppsto dyre-rekkene så plutselig i Kambrium som fossiler synes tyde på, ellre ble de langsomt splittet opp fra en felles stamform fra millioner av år tidligere? Det er ikke mulig å lage molekylær undersøkelser på fossiler fra Kambrium, men en kan sammenligne DNA-sekvenser i nålevende arter. Som nevnt ovenfor, om en går utfra en gradvis akkumulering av mutasjoner, kan en anvende antall sekvensulikheter som et molekylært ur. Formålet er å måle hvor lang tid er siden rekkene evt. ble atskilt.
Det viser seg at dateringene med en slik metode spenner vidt:Faktisk kan anslagene varierer med så mye som nærmere 100% - fra 670 millioner år siden (Russel Doolithle m.fl) til 1200 millioner år (Gregory Wray m.fl). Denne forskjellen i resultatene er for den amerikanske genetiker Kenneth Halanych tegn på at "man ikke kan tidfeste så urgamle begivenheter" når en bruker molekylære arbeidsmetoder.
For noen forskere blir valget mellom de to tidsangivelser et spørsmål om hvorvidt en velger tro på det moleklylære eller det paleontologiske forskningsmaterialet. I 1998 brukte molekylær-evolusjonistene Lindell Bromham m.fl. de molekylære data til å forkaste hypotesen om den kambriske eksplosjon. Grunnen skulle være at den beror på "en meget bokstavelig tolkning" av den fossile lagserie. I motsetning til dette uttalte paleontologene James Valentine m.fl. i 1999 at "det molekylære urs nøyaktighet er stadig problematisk, i hvert fall for oppsplittingen av rekker". Grunnen for dette var at beregningene varierte med hundrevis av millioner år, avhengig av hvilke teknikker eller molekyler en bruker. Valentine og kolleger fastholder at "de molekylære data ikke kan rokke ved den [kambriske] eksplosjonen, som stadig framstår som en hovedbegivenhet i dyrenes evolusjon.
Den kambriske eksplosjon trosser en av de betydeligste grunnsetninger i Darwins teori, ved at større dyrerekker og klasser dukker opp fra første stund. Den molekylære fylogenese løser ikke problemet, fordi de tidsangivelser som utledes av den spenner over alt for store tidsrom. Det viser seg også at dette er del av et større problem. Siden 1970-årene har evolusjonsbiologer håpet at sekvensanalyser ville overvinne mange av de vansker som de mer tradisjonelle tilnærminger har reist. Intensjonen var at en kunne sette dem i stand til å oppstille et "altomfattende livets stamtre," alene basert på molekylære analyser. Nyere oppdagelser har veltet dette håpet.
Den molekylære fylogeneses tiltagende problem
 Moderne versjoner av det darwinistiske Livets stamtre, kalles "fylogenetiske (stam)tre". På et typisk slikt tre, representerer "roten" stamformen til alle andre organismer i treet. De nederste greinene tegner de avstamningslinjer som forholdsvis tidlig har skilt seg ut, mens de høyere står for senere oppdelinger. Endene av grenene viser de egentlige arter. Oppdelingspunktet mellom to grener angir den hypotetiske stamformen for de to grenene. Mange slike trær tegnes slik at grenenes lengde svarer til forskjeller i sekvenser, som i sin tur skal indikere hvor lang tid det er gått siden de gikk hver til sitt (Fig3-7)
Moderne versjoner av det darwinistiske Livets stamtre, kalles "fylogenetiske (stam)tre". På et typisk slikt tre, representerer "roten" stamformen til alle andre organismer i treet. De nederste greinene tegner de avstamningslinjer som forholdsvis tidlig har skilt seg ut, mens de høyere står for senere oppdelinger. Endene av grenene viser de egentlige arter. Oppdelingspunktet mellom to grener angir den hypotetiske stamformen for de to grenene. Mange slike trær tegnes slik at grenenes lengde svarer til forskjeller i sekvenser, som i sin tur skal indikere hvor lang tid det er gått siden de gikk hver til sitt (Fig3-7)
Med få unntak er de eneste 'riktige' data de som svarer til grenendene på figuren. Sammensetningen av resten av treet er satt sammen ut fra metodologiske formodninger og sammenliknende sekvensanalyser. Det har vært en utbredt oppfatning om at jo flere molekyler som brukes i analysen, jo mer pålitelig må resultatet bli. Men denne forventningen begynte å smuldre opp for flere tiår siden. Molekylærbiolog James Lake m.fl. skrev i 1999 at "da forskere begynte analysere en bred vifte av gener, fra ulike organismer, fant de at organismenes innbyrdes forhold var i motstrid med rRNA-stamtreet. I følge de franske biologer Hervé Philippe og Patrick Forterre "viste det seg med flere og flere sekvensanalyser for hånden -at de fleste protein-fylogeneser er i motstrid både med hverandre og med mRNA stamtreet.
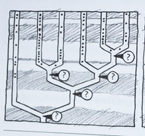 Med andre ord: ulike molekyler fører til høyst ulike fylogenetiske stamtrær. Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.
Med andre ord: ulike molekyler fører til høyst ulike fylogenetiske stamtrær. Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.
Woese tok hovedsakelig fatt på de uoverensstemmelser som befinner seg på systematikkens høyere nivåer, 'rikene'. Men problemet berører også mindre grener, herunder fylogenese for dyr. Biologen Michael Lynch skrev i 1999: "At få klarhet i de større dyrerekkenes innbyrdes forbindelser har vist seg være et problem det er vanskelig å definere nærmere. Analyser av forskjellige gener, ja selv analyser av de samme gener har brakt en mengde ulike fylogenetiske stamtrær for dagen. Selv i de tilfeller der ulike molekylanalyser kan settes sammen, er resultatet ofte bisart.
F.eks. plasserer en studie fra 1996 som har anvendt 88 proteinsekvenser, kaniner i samme gruppe som primatene i stedet for hos gnagerne. En analyser fra 1998 plasserer kua i nærmere slektskap med hvalen enn med hesten. Den molekylære fylogenesen synes være blitt plassert i en sump med mudder til halsen.
Livets stamtre rykket opp med rot
 En del molekylærbiologer mener problemet er metodologisk. I følge Forterre og Phillippe forandrer noen sekvenser seg for hurtig i evolusjonsmessig henseende til å bevare en "fylogenetisk melding" over lange tidsforløp. De hevder at ved å begrense seg til sekvenser som de mener har undergått en langsom evolusjon, kan de nå fram til et allmenngyldig stamtre. Problemet deres er at deres analyser peker i retning av en celle med kjerne som den altomfattende 'felles stamfar'. Tidligere er alltid darwinister gått ut fra at en celle uten kjerne er mer opprinnelig enn celler med kjerne. Ut fra darwinistisk synspunkt har 'treet slått rot på feil sted.'
En del molekylærbiologer mener problemet er metodologisk. I følge Forterre og Phillippe forandrer noen sekvenser seg for hurtig i evolusjonsmessig henseende til å bevare en "fylogenetisk melding" over lange tidsforløp. De hevder at ved å begrense seg til sekvenser som de mener har undergått en langsom evolusjon, kan de nå fram til et allmenngyldig stamtre. Problemet deres er at deres analyser peker i retning av en celle med kjerne som den altomfattende 'felles stamfar'. Tidligere er alltid darwinister gått ut fra at en celle uten kjerne er mer opprinnelig enn celler med kjerne. Ut fra darwinistisk synspunkt har 'treet slått rot på feil sted.'
Andre biologer, som Woese, mener at problemet ikke bare er metodologisk. Han holder fast ved at uoverensstemmelsene er tilstrekkelig hyppige og så statistisk holdbare at en verken kan late som ingenting eller bare avvise dem av metodologiske grunner. I følge ham er det på tide at en "tar de grunnleggende antakelser opp til gjenoverveielse." Han anbefaler at en forlater idéen om en felles organisme som stamfar. Alternativet han stiller opp som grunnleggende er "en prosess". Men om den 'altomfattende felles stamfar' ikke er en organisme, gir det vel heller liten mening i å snakke om en 'felles forfader'.
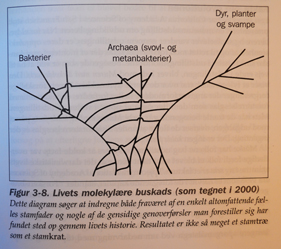 En annen løsning på problemet er framsatt av W.Ford Doolittle, biolog ved Dalousie University. Han skriver i 1999: "Det kan godt være at det ennå ikke er lykkes .. å finne det riktige tre,..fordi livets historie i bunn og grunn ikke kan framstilles som et tre." I følge Doolittle skyldes de fylogenetiske uoverensstemmelser i overveiende grad "en gjensidig genoverføring." Doolittle lanserer den idé at genoverførsel mellom bakterier og urceller seg i mellom ligger til grunn for mange av uoverensstemmelsene i molekylære fylogeneser. Det innebærer at livets tidlige historie ikke ville likne et tre, men mer ett sammenflettet kratt (Fig3-8)
En annen løsning på problemet er framsatt av W.Ford Doolittle, biolog ved Dalousie University. Han skriver i 1999: "Det kan godt være at det ennå ikke er lykkes .. å finne det riktige tre,..fordi livets historie i bunn og grunn ikke kan framstilles som et tre." I følge Doolittle skyldes de fylogenetiske uoverensstemmelser i overveiende grad "en gjensidig genoverføring." Doolittle lanserer den idé at genoverførsel mellom bakterier og urceller seg i mellom ligger til grunn for mange av uoverensstemmelsene i molekylære fylogeneser. Det innebærer at livets tidlige historie ikke ville likne et tre, men mer ett sammenflettet kratt (Fig3-8)
Doolittle uttaler: "kanskje det ville være bedre, og i det lange løp mer produktivt, å gå vekk fra forsøket på å tvinge dataene .. inn i den form Darwin har skapt." I en artikkel fra februar år 2000 i Scientific American (Uprooting the tree of life) nådde han samme konklusjon: " Vi har bruk for nye hypoteser, som vil munne ut i noe vi ikke engang tør gjette på hva er." Så mønsteret i det forgrenede  evolusjonstreet stemmer ikke med de større trekk i det fossile og molekylære vitnesbyrd. Den kambriske eksplosjon viser at de høyeste kategorier er oppstått først, og det vender opp ned på Darwins evolusjonstre. (Se mer i "Darwins Doubt lenger ned i bloggen") Allikevel har "Livets Stamtre" stadig en dominerende plass blant evolusjonens ikoner, fordi darwinister har utropt det til å være et faktum!
evolusjonstreet stemmer ikke med de større trekk i det fossile og molekylære vitnesbyrd. Den kambriske eksplosjon viser at de høyeste kategorier er oppstått først, og det vender opp ned på Darwins evolusjonstre. (Se mer i "Darwins Doubt lenger ned i bloggen") Allikevel har "Livets Stamtre" stadig en dominerende plass blant evolusjonens ikoner, fordi darwinister har utropt det til å være et faktum!
Til tross for de nyere oppdagelsene innen molekylær fylogenese, holder National Academy of Scienses bastant fast ved at "molekylærbiologiens vitnesbyrd om evolusjonen som overveldende, og det vokser til stadighet" Det virker som de ikke holder følge med den forskning de selv skulle lede an i. Det blir da ikke så rart at lærebøker i biologi ikke følger opp, selv om 100 års-søvn er alt for lenge (se Haeckels fostre i min blogg). De tviler ikke et sekund på 'det overveldende vitnesbyrd i Livets Stamtre er en realitet, og at ingen forsker så mye som ville drømme om å tvile på en altomfattende felles avstamning."
Fakta om evolusjonen
Harry Whittington, som var paleontologen som først åpenbarte omfanget av den kambriske eksplosjonen, skrev i 1985: "Jeg ser med stor skepsis på de diagrammer som viser at dyrelivets diversitet skulle være resultat av en forgrening opp gjennom tiden, og som koker det ned til avstamning fra en eller annen form for dyr. . Dyrene kan meget vel være  oppstått mer enn én gang og på ulike steder til ulike tider." Etter å ha fått tilgang til nyeste informasjon om molekylær fylogenese, skriver biologen Malcolm Gordon i 1998: "livet ser ut til å ha hatt atskillige opprinnelseskilder. Det universelle livets stamtre synes å ha mer enn bare en rot.. Den tradisjonelle versjon av teorien om felles avstamning, gjelder tilsynelatende ikke for riker ..[eller] rekker, og muligens heller ikke for mange av klassene innenfor rekkene.
oppstått mer enn én gang og på ulike steder til ulike tider." Etter å ha fått tilgang til nyeste informasjon om molekylær fylogenese, skriver biologen Malcolm Gordon i 1998: "livet ser ut til å ha hatt atskillige opprinnelseskilder. Det universelle livets stamtre synes å ha mer enn bare en rot.. Den tradisjonelle versjon av teorien om felles avstamning, gjelder tilsynelatende ikke for riker ..[eller] rekker, og muligens heller ikke for mange av klassene innenfor rekkene.
Kompetente biologer setter spørsmålstegn ved det darwinistiske Livets Stamtre. Bedømt ut fra det fossile og molekylære vitnesbyrd, har vi her ikke å gjøre med en velbegrunnet teori. Men om en reiser spørsmål ved det, blir det vanligvis ikke sett på med milde øyne. Særlig gjelder det kanskje USA, men også her i Norge kan en merke motstanden, -også på kirkelig hold. Det er fullt tillatt å kritisere regjering og styrende, men å kritisere Darwin blir ikke vel mottatt.
Utvalg av stoff og bilder ved Asbjørn E. Lund