Topoisomerase II enzymer, fantastisk bevis på design
 Fullstendige og like overføringer av DNA til datterceller er avgjørende under mitose. Under celledeling arver hver dattercelle en kopi av hvert kromosom. Overgangen metafase-til-anafase er det kritiske punktet i cellesyklusen der cellen forplikter seg til separasjon av søster-kromatider. Når spindelfesteret er fullført, må kohesjon fjernes for å muliggjøre fysisk separasjon av søsterkromatider. Dette krever spaltning av proteinkomplekset kohesin ved separase og i noen tilfeller fullføring av kromosom-dekatenasjon. Katenering er prosessen der to sirkulære DNA-tråder er koblet sammen som kjedeledd. Dette skjer etter DNA-replikasjon, der to enkeltstrenger er katenert og fortsatt kan replikere, men ikke kan skilles ut i de to dattercellene.
Fullstendige og like overføringer av DNA til datterceller er avgjørende under mitose. Under celledeling arver hver dattercelle en kopi av hvert kromosom. Overgangen metafase-til-anafase er det kritiske punktet i cellesyklusen der cellen forplikter seg til separasjon av søster-kromatider. Når spindelfesteret er fullført, må kohesjon fjernes for å muliggjøre fysisk separasjon av søsterkromatider. Dette krever spaltning av proteinkomplekset kohesin ved separase og i noen tilfeller fullføring av kromosom-dekatenasjon. Katenering er prosessen der to sirkulære DNA-tråder er koblet sammen som kjedeledd. Dette skjer etter DNA-replikasjon, der to enkeltstrenger er katenert og fortsatt kan replikere, men ikke kan skilles ut i de to dattercellene.
II Topoisomerase-enzymer er et allestedsnærværende enzym som er essensielt for overlevelse hos alle eukaryote organismer og spiller kritiske roller i praktisk talt alle aspekter av DNA-metabolismen. Den utfører den fantastiske bragden å bryte en DNA-helix, føre en annen helix gjennom spalten og forsegle den doble helixen bak den. De er viktige for separasjon av sammenfiltrede datterstrenger under replikering. Denne funksjonen antas å bli utført av topoisomerase II i eukaryoter og av topoisomerase IV i prokaryoter. Unnlatelse av å skille disse strengene fører til celledød. Som genetisk materiale er DNA fantastisk, men som makromolekyl er det ustyrlig, voluminøst og skjørt. Uten virkningen av DNA-replikaser, topoisomeraser, helikaser, translokaser og rekombinaser, ville genomet kollapse i en topologisk sammenfiltret tilfeldig spole som ville være ubrukelig for cellen. Topoisomerasen antas å være en svært dynamisk struktur, med flere porter for innføring av DNA i de to DNA-størrelse hullene. Tap av topoisomeraseaktivitet i metafase fører til forsinket utgang og omfattende anafase kromosom-bridging, noe som ofte resulterer i cytokinesis-svikt, selv om opprettholdelse av begrenset katenasjon inntil anafase kan være viktig for søster-kromatidens strukturelle organisering. (9) Nøyaktig overføring av kromosomer krever at søsterens DNA-molekyler opprettet under DNA-replikasjon skilles fra og deretter trekkes til motsatte poler i cellen før deling. Defekter ved kromosom segregasjon produserer celler som er aneuploide (som inneholder et unormalt antall kromosomer) - en situasjon som kan få alvorlige konsekvenser.
Som mange andre enzymer er topoisomerase II essensiell for cellefunksjon, og måtte være til stede i den første levende cellen for å utøve sin funksjon helt i begynnelsen, da livet begynte.
Innenfor hvert kromosom spiller to dimensjoner av organisasjonen: kondensering langs aksene sikrer at hele kromatid, fra ende til ende, holdes sammen 8, mens den tette foreningen av søster-kromatider inntil anafase, betegnet SøsterKromatid-Kohesjon (SKK), som sikrer at hver dattercelle bare mottar en kopi. Det er kjent at to mekanismer spiller en rolle i SKK: DNA-katenering, som fysisk sperrer (katenerer) DNA på tvers av søsterkromatidene; og proteinkoblinger gjennom kohesinkomplekset, som fysisk binder søsterkromatidene til hverandre.
{Tillegg: For de som er visuelt anlagt, anbefales denne videoen -Topoisomerase (4m:45s). Lagt til av oversetter}
Topoisomerase II danner en kovalent kobling til begge strengene av DNA-heliksen samtidig, noe som gjør en kortvarig dobbeltstrengbrudd i helixen. Disse enzymene aktiveres av steder på kromosomer der to doble helikser krysser hverandre, slik som de som genereres ved supercoiling foran en replikasjonsgaffel
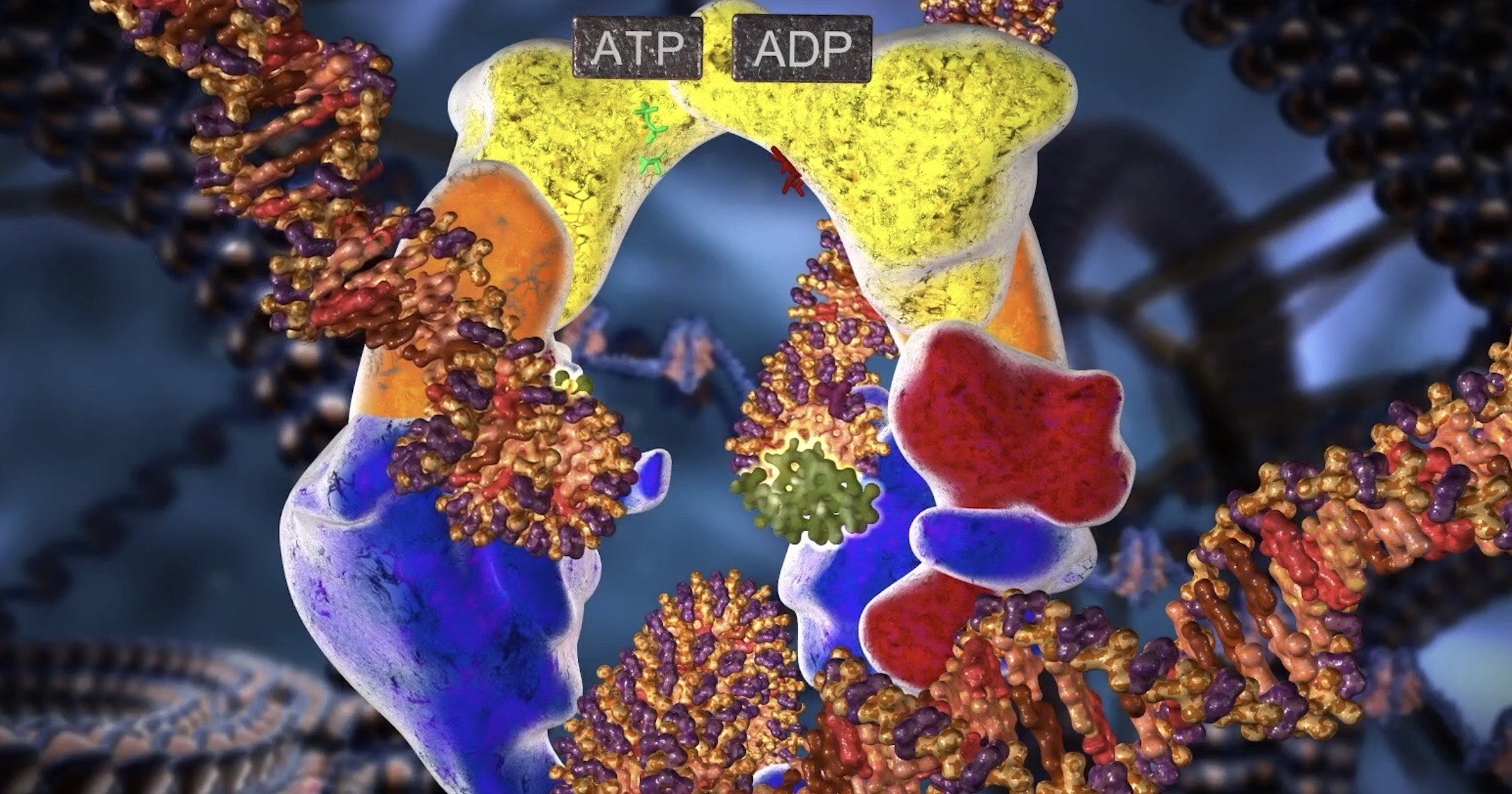 Når et topoisomerase II-molekyl binder seg til et slikt kryssingssted, bruker proteinet ATP-hydrolyse for å utføre følgende reaksjoner effektivt:
Når et topoisomerase II-molekyl binder seg til et slikt kryssingssted, bruker proteinet ATP-hydrolyse for å utføre følgende reaksjoner effektivt:
(1) den bryter en dobbelt helix reversibelt for å lage en DNA "gate";
(2) det fører til at den andre, nærliggende dobbeltspiralen passerer gjennom denne åpningen; og
(3) den lukker deretter bruddet og dissosierer seg fra DNAet. Ved overgangspunkter generert av supercoiling skjer passering av dobbelt helix gjennom porten i retningen som vil redusere supercoiling. På denne måten kan type II topoisomeraser avlaste overvind-spenningen som genereres foran en replikasjonsgaffel. Deres reaksjonsmekanisme lar også DNA-topoisomeraser av type II effektivt skille to sammenlåste DNA-sirkler. Topoisomerase II forhindrer også alvorlige DNA-sammenfiltringsproblemer som ellers ville oppstå under DNA-replikasjon. Topoisomerase IIs enorme nytteverdi for å fjerne kromosomer kan lett forstås av alle som har kjempet for å fjerne en floke fra et fiskesnøre uten hjelp av saks/kniv.
Disse molekylære maskinene er langt utenfor hva ikke-styrte prosesser som involverer sjanse og nødvendighet kan produsere. Faktisk er maskiner med kompleksitet og sofistikering av Topoisomerase-enzymer, basert på vår erfaring, vanligvis tilskrevet intelligente agenter.
 Topoisomeraser av type IIA består av flere nøkkelmotiver:
Topoisomeraser av type IIA består av flere nøkkelmotiver:
ett N-terminalt GHKL ATPase-domenee
Toprim-domene
sentral DNA-bindende kjerne
C-terminalt domene
Hvert av disse nøkkelmotivene er viktige for at enzymet skal fungere riktig. Ingen deler kan reduseres, og det er heller ikke mulig at noen av under-delene dukker opp med naturlige midler. Ikke bare hadde enzymet dukket opp før den første cellen ble dannet, og det kunne derfor ikke være et resultat av evolusjon, men underdelene i seg selv, og enzymet i seg selv til og med fullstendig dannet, ville ikke ha noen nytte, med mindre DNA dobbelt helix molekyler allerede også var eksisterende, og dermed hele prosessen med celledeling, mitose og katenasjon, som skjer gjennom DNA-replikasjon. Enzymet er imidlertid essensielt for livet, så hvis Topo II fjernes, kunne ikke livet eksistere. Så vi har her en av utallige essensielle tilsynelatende bittesmå og aparte uviktige deler, som ved nærmere ettersyn viser seg være livsviktige. Dette gir et annet stort spørsmålstegn når det gjelder naturalistiske forklaringer, men gir på den andre siden et mer kraftig argument for design.
Oversettelse og bilder ved Asbjørn E. Lund