Boolsk algebra i celler
Oversatt fra Angelo Grasso (FB), omtales her. (kursiv og understreking ved oversetter)
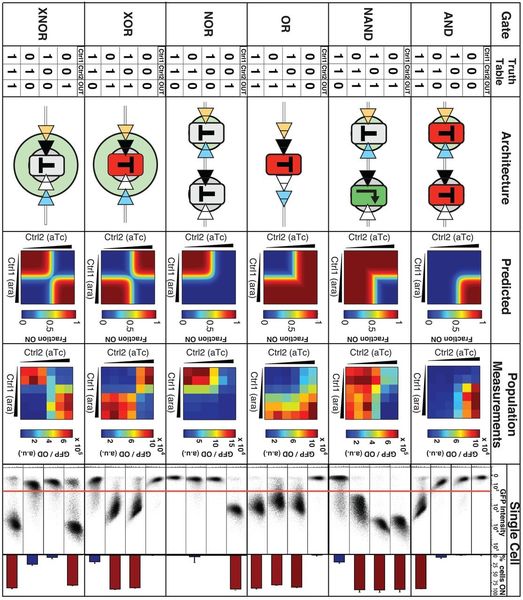 1. Celler behandler kontinuerlig mangfoldet av inngangssignaler for å ta avgjørelser om passende svar som fører til endringer i genuttrykk, enzymatisk aktivitet, omlegging av signalnettverk, migrasjon, vekst eller deling, samt programmert celledød som utdata informasjon. I en beregningsenhet behandles inngangsinformasjonen matematisk til et digitalt signal. Dette signalet er en kode som representerer de fysiske signalene og antar en sekvens av diskrete verdier. For eksempel, når det gjelder en binær kode, blir den grunnleggende informasjonsenheten betegnet som en serie med '0' og '1' sifre. De binære sifrene indikerer de to tilstandene i den logiske kretsen. En terskel er implementert for å definere inngangs- og utgangsområdet som kan kategoriseres under hvert logikksett. Hvis verdien enten er lavere eller høyere enn terskelen, er tilstanden til kretsen definert som henholdsvis '0' eller '1'. Digitale kretser bruker omfattende logiske elementer som er sammenkoblet for å skape logiske porter, som er i stand til å utføre boolske logiske funksjoner, inkludert NOT, OR, AND, og alle deres mulige kombinasjoner.
1. Celler behandler kontinuerlig mangfoldet av inngangssignaler for å ta avgjørelser om passende svar som fører til endringer i genuttrykk, enzymatisk aktivitet, omlegging av signalnettverk, migrasjon, vekst eller deling, samt programmert celledød som utdata informasjon. I en beregningsenhet behandles inngangsinformasjonen matematisk til et digitalt signal. Dette signalet er en kode som representerer de fysiske signalene og antar en sekvens av diskrete verdier. For eksempel, når det gjelder en binær kode, blir den grunnleggende informasjonsenheten betegnet som en serie med '0' og '1' sifre. De binære sifrene indikerer de to tilstandene i den logiske kretsen. En terskel er implementert for å definere inngangs- og utgangsområdet som kan kategoriseres under hvert logikksett. Hvis verdien enten er lavere eller høyere enn terskelen, er tilstanden til kretsen definert som henholdsvis '0' eller '1'. Digitale kretser bruker omfattende logiske elementer som er sammenkoblet for å skape logiske porter, som er i stand til å utføre boolske logiske funksjoner, inkludert NOT, OR, AND, og alle deres mulige kombinasjoner.
2. Molekylære og biomolekylære logiske porter og deres nettverk behandler kjemiske inngangssignaler på lignende vis som på menneskeskapte datamaskiner. Likhetene i behandlingen av informasjon av biologiske systemer og menneskelig designede enheter er bredt anerkjent av mange forskere.
3. Beslutningstaking er enten a) noe som utføres direkte av intelligens, eller b) programmert av intelligens som skal utføres av maskiner, som datamaskiner. Det har aldri blitt observert at ikke-ledede evolusjonære mekanismer kunne produsere en beslutningstaker med det formål å generere spesifikke resultater lenger fremme. Fremsyn er en kvalitet utelukkende utført av intelligens. Hvis analogien til to fenomener er veldig nær og slående, mens årsaken til det ene samtidig er veldig åpenbar, blir det knapt mulig å nekte å innrømme handlingen av en analog årsak i den andre, men ikke så åpenbar i seg selv. En metafor ("En biologisk celle er som et produksjonssystem") viser at lignende oppførsel drives av lignende årsaksmekanismer.
4. Derfor er beslutningsprosessen observert i cellen sannsynligvis et resultat av guddommelig superintelligent design.
Bakterier bruker DNA og proteiner for å implementere en if /else -uttalelse for å regulere glukosenivåer ved hjelp av boolsk logikk ('Lac operon') (8)
Gener utfører rutinemessig betinget "hvis-så" -logikk (alle gener aktiveres bare hvis spesifikke forhold er oppfylt), "utfør-løkker" (visse gener skaper et bestemt antall kroppssegmenter og deler; for eksempel skade på et gen som kalles lunefullt Sonic Hedgehog kan produsere ekstra fingre), 'timingrutiner' (gener koder for klokkeproteiner), 'subrutineanrop' (et gen kalt Pax6 i bananfluer kan initiere en genkaskade som rekrutterer over 2000 spesialiserte gener for å bygge forskjellige deler av øyet) og så videre.
Når programvareingeniører jobber med komplekse programmeringsprosjekter, finner de det fordelaktig å organisere koden i moduler, slik at alle rutinene som er ansvarlige for en bestemt del av applikasjonen, er i samme modul. De kan da lett nås for spesifikke oppgaver knyttet til den felles funksjonen. Antagelig er den samme fordelen for cellen. Ved å ha alle genene som er ansvarlige for en bestemt funksjon, for eksempel hankjønn, på det samme kromosomet, kan alle slike gener lettere kontrolleres av genpromotorer og -hemmere som er ansvarlige for mannlige egenskaper. (7)
Binær logikk eller boolsk logikk er kjernekonseptet med boolsk algebra som danner 'kanaler' som alle digitale elektroniske kretser og mikroprosessorbaserte systemer er konstruert av. Grunnleggende digitale logikk-porter utfører logiske operasjoner av AND, OR og NOT på binære tall. Informasjon lagres i datasystemer i binær form. En binær bit representerer en av de to mulige tilstandene, som vanligvis refereres til som logisk '1' og logisk '0'. Nærmere bestemt kan tilstedeværelsen av en positiv spenning representeres som logisk "1", høy eller sann; fraværet av en spenning kan representeres som logisk "0", lav eller usann. I boolsk algebra og sannhetstabell er disse to tilstandene representert som henholdsvis '1' og '0'.
 Oppkalt etter George Boole, en matematiker fra 1800-tallet, er boolsk logikk grunnlaget for digital kretsdesign. AND-funksjonen betyr for eksempel at begge betingelsene må være på, for å utløse kretsen. I notasjonsform, der 1 betyr til stede og 0 betyr fraværende,
Oppkalt etter George Boole, en matematiker fra 1800-tallet, er boolsk logikk grunnlaget for digital kretsdesign. AND-funksjonen betyr for eksempel at begge betingelsene må være på, for å utløse kretsen. I notasjonsform, der 1 betyr til stede og 0 betyr fraværende,
1 OG 1 = sann eller 'på'
0 OG 0 = falsk eller 'av'
1 OG 0 = falsk eller 'av'
I ELLER-funksjonen kan enten en (eller begge) av inngangene være på for å trigge kretsen:
1 ELLER 1 = sann eller 'på'
0 ELLER 0 = falsk eller 'av'
1 ELLER 0 = sant eller 'på'
Negasjonen av disse funksjonene, kalt NAND og NOR, reverserer utgangene. For en logisk kanal med to innganger og en utgang, for eksempel en transistor, er det 16 mulige operasjoner (AND, NAND, OR, NOR, XOR, XNOR, ..
Probabilistiske boolske nettverk: modellering og kontroll av genregulerende nettverk, Ilya Shmulevich, Edward R. Dougherty, side 12: En av de enkleste representasjonsrammene antar at gener er binære verdier, noe som indikerer at de kan være i én av to mulige tilstander av aktivitet (f.eks. På eller AV) til et gitt tidspunkt, og at de virker på hverandre ved hjelp av regler representert ved boolske funksjoner.
Celler behandler kontinuerlig dette mangfoldet av inngangssignaler for å ta avgjørelser om passende svar som fører til endringer i genuttrykk, enzymatisk aktivitet og omlegging av signalnettverket.
Grassos kommentar: Beslutningstaking er enten a) noe som utføres direkte av intelligens, eller b) er programmert av intelligens som skal utføres av maskiner, som datamaskiner. Det har aldri blitt observert at ikke-styrte evolusjonære mekanismer kunne produsere en beslutningstakings maskin med det formål å generere spesifikke resultater lenger fremme. Fremsyn er en kvalitet utelukkende utført av intelligens.
Denne beslutningsprosessen manifesterer seg i form av migrasjon, vekst eller deling, samt programmert celledød som utdatainformasjonen. I en beregningsenhet behandles inngangsinformasjonen matematisk til et digitalt signal. Dette signalet er en kode som representerer de fysiske signalene og antar en sekvens av diskrete verdier. For eksempel, når det gjelder en binær kode, blir den grunnleggende informasjonsenheten betegnet som en serie med '0' og '1' sifre. De binære sifrene indikerer de to tilstandene i den logiske kretsen. En terskel er implementert for å definere inngangs- og utgangsområdet som kan kategoriseres under hvert logikksett. Hvis verdien enten er lavere eller høyere enn terskelen, er tilstanden til kretsen definert som henholdsvis '0' eller '1'. Digitale kretser bruker omfattende logiske elementer som er sammenkoblet for å skape logiske porter, som er i stand til å utføre boolske logiske funksjoner, inkludert NOT, OR, AND, og alle deres mulige kombinasjoner (5)
Molekylære og biomolekylære logiske porter og deres nettverk behandler kjemiske inngangssignaler på lignende vis som datamaskiner. Likhetene i behandlingen av informasjon av biologiske systemer og menneskelig designede enheter er bredt anerkjent av mange forskere.
Grassos kommentar: Herschel 1830 1987, s. 148:
"Hvis analogien mellom to fenomener er veldig nær og slående, mens årsaken til det ene på samme tid er veldig åpenbar, blir det knapt mulig å nekte å innrømme handlingen av en analog årsak i den andre, om ikke så åpenbar i seg selv."
En metafor ("En biologisk celle er som et produksjonssystem") viser at lignende oppførsel drives av lignende årsaksmekanismer.
Digitale kretser bruker omfattende logiske elementer som er sammenkoblet for å lage logiske porter, som er i stand til å utføre boolske logiske funksjoner, inkludert NOT, OR, AND og alle deres mulige kombinasjoner (3)
Genomisk kontrollprosess: utvikling og evolusjon av Isabelle S. Peter, Eric H. Davidson side 45
Dyrekropper viser en grunnleggende boolsk karakter på alle nivåer i organisasjonen. Kroppsdelene som de består av, er diskrete, og deres struktur og funksjon samt plassering i kroppen er deterministisk programmert og romlig avgrenset.
 Evolusjonær biovitenskap som reguleringssystembiologi [2011] Davidson, Eric (4)
Evolusjonær biovitenskap som reguleringssystembiologi [2011] Davidson, Eric (4)
Hierarkiet og flere tilleggsdesignfunksjoner i disse nettverkene virker til å produsere boolske regulatoriske tilstands-spesifikasjonsfunksjoner i oppstrømsfaser av utviklingen av kroppsplanen. Uansett kontinuerlig variasjon som forekommer ved individuelle cis-regulatoriske sekvenser, bevarer dGRN-kretsutgangen sin boolske morfogenetiske karakter.
dGRN-kretsløp styrer romlig reguleringstilstand under utvikling som utfører boolske logiske transaksjoner.
Boolsk nettverksdynamikk kan brukes til å forstå cellulære tilstander som spredning, differensiering og apoptose. (2)
Evolusjonær biovitenskap som reguleringssystembiologi 2011 Eric H. Davidson
Akkurat som utvikling er en systemegenskap for det regulatoriske genomet, må det vurderes på systemnivå at utviklingsprosessen kan endre seg. Endring av kroppsplanen avhenger av endring av strukturen i utviklingsgenereguleringsnettverk som helhet. Hierarkiet og flere tilleggsdesignfunksjoner i disse nettverkene virker for å produsere boolske regulatoriske tilstandsspesifikasjonsfunksjoner i oppstrømsfaser av utviklingen av kroppsplanen. Disse er skapt av logiske utganger fra nettverks-underkretser, og hos moderne dyr er disse utgangene ugjennomtrengelige for kontinuerlig adaptiv variasjon.
Dyrekroppsplaner er en systemnivåegenskap for utviklingsgenereguleringsnettverk (dGRNs) som styrer ontogeni i kroppsplanen. Det følger at store morfologisk nyheter krever dramatiske endringer i dGRN-arkitekturen, som alltid involverer flere regulatoriske gener, og som typisk påvirker distribusjonen av hele nettverks-underkretser. Fordi dGRN er dypt hierarkiske, og det er de øvre nivåene av disse GRNene som styrer de viktigste morfologiske funksjonene i utviklingen, oppstår et spørsmål som behandles nedenfor i dette essayet: Hvordan kan vi tenke på naturlig utvalg i forhold til dGRN-organisering? Svarene ligger i dGRNs arkitektur og utviklingslogikken de genererer på systemnivå, langt fra mikroevolusjonær mekanisme. Mens adaptiv evolusjonær variasjon forekommer konstant hos moderne dyr i periferien av dGRNs, krever stabiliteten over geologiske epoker av utviklingsegenskapene som definerer de viktigste egenskapene til kroppsplanene deres, spesielle forklaringer forankret dypt i struktur/funksjonsforholdene hos dGRNs. (6)
Spørsmål: Hva er disse spesielle forklaringene?
Oversettelse og bilder ved Asbjørn E. Lund