Rask evolusjon skjer i dag
(oversatt fra 'The Evolution Revolution' Kap. 3 -av Dr. Lee M. Spetner, Juni 2014.)
 I dette kapitlet vil vi se på flere representative eksempler på at evolusjon skjer i våre dager. Noen av eks. kan forklares ved en Ikke-Tilfeldig Evolusjonshypotese, som vi har sett på tidligere. Bare ved direkte observasjoner, som disse eksemplene kan en håpe å forstå hvordan evolusjon og i hvilken utstrekning, om i det hele tatt, begrepet kan utvides til å gjelde i forhold til felles avstamning. Evolusjon i betydningen endring i populasjoner forekommer, men det er dårlig vitenskap å slutte fra slike eks. at felles avstamning er en realitet. Det vil bli klarere i løpet av eksemplene.
I dette kapitlet vil vi se på flere representative eksempler på at evolusjon skjer i våre dager. Noen av eks. kan forklares ved en Ikke-Tilfeldig Evolusjonshypotese, som vi har sett på tidligere. Bare ved direkte observasjoner, som disse eksemplene kan en håpe å forstå hvordan evolusjon og i hvilken utstrekning, om i det hele tatt, begrepet kan utvides til å gjelde i forhold til felles avstamning. Evolusjon i betydningen endring i populasjoner forekommer, men det er dårlig vitenskap å slutte fra slike eks. at felles avstamning er en realitet. Det vil bli klarere i løpet av eksemplene.
Tusenfryd har frø som kan spres med vinden. Hvert lite frø er omgitt av en mye større luftig dun, som gir den stor overflate og tilfører lite ekstra vekt. Slike vindbasert spredning av plantens DNA er oftest en fordel, men det finne eks. på det motsatte. På små øyer ville tusenfrydens vindbaserte spredning kunne resultere i at de fleste frø gikk tapt. For å imøtekomme denne muligheten utvikler tusenfryd seg til å stoppe langdistanse-spredning av frø, når det ble en ulempe. M.Cody og J.Overton (tidligere) UCLA studerte mer enn 200 øyer av ulike størrelse som hadde tusenfryd-liknende planter. De oppdaget at når disse typer planter ble fraktet fra fastlandet til små øyer, tapte de sin evne til langdistanse-frøspredning bare i løpet av noen få år. Disse endringene er genetisk baserte og arvelige.
Bilde 1. Tusenfryd frø kan også spres av vind
Dette ligner en Ikke-tilfeldig evolusjon , der endringer i omgivelsene trigger en epigenetiske endring, som er tilpasset de nye omgivelsene. Det kan også hevdes at genetisk konfigurasjon for tap av evne til frøspredning alltid hadde eksistert i populasjonen, og når forholdene favoriserer det, kommer det til sin rett. Uansett blir det ikke tilført noen ny informasjon, til genomet i tusenfryd. I det ene tilfellet trenger det bare å trigges fra omgivelsene. I det andre tilfellet er det to distinkte genom i populasjonen, men begge var der fra start av. Selv om eks. tjener som bilde på evolusjon i en populasjon, så gir det intet tilskudd til dogmet om felles avstamning.
 Eple-larve fluen Rhagoletis momonella spiste og utbredte seg opprinnelig i treet hagetorn, men i det 19. århundre forlot flua det og begynte å infisere epletre. Den har nå spredt seg til å ernære og forplante seg også på roser, pærer og kirsebær. Da fluene antok en ny vert, så gjennomgikk de flere atferdsmodifikasjoner: Deres preferanse endret seg fra den gamle til den nye verten. De adopterte nye parringspreferanser som isolerte de fra den gamle populasjonen. Dette hindret dem i å blande seg. De tilpasset modningstid til modning av den nye vertsfrukten. Alle disse endringene har vist seg å være gentisk basert og er arvelige (Barton et al. 1988, Feder et al. 1988, McPherong et al 1988, Smith 1988). Det krever da endringer i genomet for å framvise endringer. Alle disse endringene måtte skje raskt og samtidig, noe en sekvens av tilfeldige feil i DNA-replikasjon ikke kan avstedkomme. Men en innebygd tilsvars-mulighet lik den som er innebygd i en Ikke-tilfeldig evolusjonsmekanisme kan utføre det. Det er heller ikke her noen økning i informasjon, som kan rettferdiggjøre å benytte eks. i en sammenheng om felles avstamning.
Eple-larve fluen Rhagoletis momonella spiste og utbredte seg opprinnelig i treet hagetorn, men i det 19. århundre forlot flua det og begynte å infisere epletre. Den har nå spredt seg til å ernære og forplante seg også på roser, pærer og kirsebær. Da fluene antok en ny vert, så gjennomgikk de flere atferdsmodifikasjoner: Deres preferanse endret seg fra den gamle til den nye verten. De adopterte nye parringspreferanser som isolerte de fra den gamle populasjonen. Dette hindret dem i å blande seg. De tilpasset modningstid til modning av den nye vertsfrukten. Alle disse endringene har vist seg å være gentisk basert og er arvelige (Barton et al. 1988, Feder et al. 1988, McPherong et al 1988, Smith 1988). Det krever da endringer i genomet for å framvise endringer. Alle disse endringene måtte skje raskt og samtidig, noe en sekvens av tilfeldige feil i DNA-replikasjon ikke kan avstedkomme. Men en innebygd tilsvars-mulighet lik den som er innebygd i en Ikke-tilfeldig evolusjonsmekanisme kan utføre det. Det er heller ikke her noen økning i informasjon, som kan rettferdiggjøre å benytte eks. i en sammenheng om felles avstamning.
Bilde 2. Eplefluen Rhagoletis momonella
 Nesten alle med et lite akvarium hjemme, er kjent med guppier. Det har vært rapportert at disse fiskene framviser rask evolusjon (Reznick et al. 1990, Carrol et al. 2007, Gordon et al. 2009). Disse små fiskene jaktes på av større rovfisk. Ciclide-fisk jakter på større, modne guppier, mens karper jakter på de mindre. Det viser seg at guppier tilpasser sin morfologi og oppførsel etter type rovfisk i deres område. I nærheten av ciklider, tilpasser guppier seg ved å modnes tidlig og ha mange små avkom, noe som minker fare for å bli tatt av ciklider. I nærheten av karper, tenderer guppier til å modnes sent og ha færre, men større avkom, som gjør at færre bli bytte for karpene.
Nesten alle med et lite akvarium hjemme, er kjent med guppier. Det har vært rapportert at disse fiskene framviser rask evolusjon (Reznick et al. 1990, Carrol et al. 2007, Gordon et al. 2009). Disse små fiskene jaktes på av større rovfisk. Ciclide-fisk jakter på større, modne guppier, mens karper jakter på de mindre. Det viser seg at guppier tilpasser sin morfologi og oppførsel etter type rovfisk i deres område. I nærheten av ciklider, tilpasser guppier seg ved å modnes tidlig og ha mange små avkom, noe som minker fare for å bli tatt av ciklider. I nærheten av karper, tenderer guppier til å modnes sent og ha færre, men større avkom, som gjør at færre bli bytte for karpene.
For å studere hvordan disse tilpasningene inntraff, så har D. Reznick og al. studert guppier i Trinidad (Aripo elven). Her forekommer ciklider sammen med guppier. En sideelv har karper, men ingen ciklider eller guppier, inntil Reznick og al. oveførte 200 guppier fra hovedelva. Snart dukket det opp endringer i guppy-populasjonen. Det endret seg til det som er vanlig i forhold til karpe, og Reznick fant at endringene var arvelige. De første endringene ble observert etter bare to år, som er alt for raskt for at tilfeldige mutasjoner kan ha noen effekt. Reznick fortolket disse endringene til å kunne relateres til naturlig seleksjon, og de viste seg være arvelige. Heller ikke her ble ny informasjon dannet, og kan dermed ikke begrunne felles avstamning. Det er mulig at endringene ble generert av genom-endringer iverksatt av omgivelsene, som i en Ikke-tilfeldig evolusjonsmekanisme.
Bilde 3. Guppy -kjent fra akvarier og i naturen
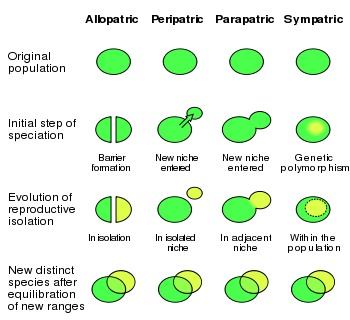 Øgler er blitt observert å tilpasse seg nye omgivelser etter bare 10-14 år (Losos, 2001), Case 1997), som er alt for raskt for tilfeldige mutasjonsendringer å bli selektert på. Øgler ble introdusert på flere øyer i Bahamas (Schoener et al. 1983). Etter ca. 10 år ble det oppdaget at de hadde utviklet seg i ulike retninger. De hadde tilpasset seg ulike nisjer og viste således rask artsdannelse. Endringene viste et parallelt forløp som på de fire øyene i Antillene: Puerto Rica, Cuba, Hispanolia og Jamaica. Konvergent og parallell evolusjon er høyst usannsynlig som følge av tilfeldige mutasjoner (Spetner 1997). Denne type endringer kunne imidlertid forventes ut fra at genetiske endringer ble trigget ut fra omgivelsesendringer. Dette synes å være artsdannelse i samme eller overlappende geografiske område og populasjoner (sympatric speciation), heller enn i atskilte områder og så kommet sammen senere (allopatric speciation). Etter 20 år ble hver øy funnet å ha flere atskilte øgle-arter, som skilte seg fra den opprinnelig importerte øgle-arten. DNA-analyser viste at utvikling skjedde der på øya, og ikke ble immigrert fra omgivelsene. Det ble vist ved at øgle-DNA fra ulike øgle-arter på samme øy viste større samsvar enn mer like øgle-arter på andre øyer. (Losos, 2001, Jackman et al. 1997). Innebygde mekanismer i øglene var latent til stede, og ble trigget av endriger i omgivelsene.
Øgler er blitt observert å tilpasse seg nye omgivelser etter bare 10-14 år (Losos, 2001), Case 1997), som er alt for raskt for tilfeldige mutasjonsendringer å bli selektert på. Øgler ble introdusert på flere øyer i Bahamas (Schoener et al. 1983). Etter ca. 10 år ble det oppdaget at de hadde utviklet seg i ulike retninger. De hadde tilpasset seg ulike nisjer og viste således rask artsdannelse. Endringene viste et parallelt forløp som på de fire øyene i Antillene: Puerto Rica, Cuba, Hispanolia og Jamaica. Konvergent og parallell evolusjon er høyst usannsynlig som følge av tilfeldige mutasjoner (Spetner 1997). Denne type endringer kunne imidlertid forventes ut fra at genetiske endringer ble trigget ut fra omgivelsesendringer. Dette synes å være artsdannelse i samme eller overlappende geografiske område og populasjoner (sympatric speciation), heller enn i atskilte områder og så kommet sammen senere (allopatric speciation). Etter 20 år ble hver øy funnet å ha flere atskilte øgle-arter, som skilte seg fra den opprinnelig importerte øgle-arten. DNA-analyser viste at utvikling skjedde der på øya, og ikke ble immigrert fra omgivelsene. Det ble vist ved at øgle-DNA fra ulike øgle-arter på samme øy viste større samsvar enn mer like øgle-arter på andre øyer. (Losos, 2001, Jackman et al. 1997). Innebygde mekanismer i øglene var latent til stede, og ble trigget av endriger i omgivelsene.
Bilde 4. Ulike typer artsdannelse
Evolusjon av Darwins finker
 Som kjente besøkte Ch. Darwin Galapagos under sin tur med H.M.S. Beagle. På øyene fant han finker som ikke så ut som finkene han hadde sett tidligere. Ekspertstudier av kolleger i London bekreftet dem å være nye finke-arter. Darwin teoretiserte at for lenge siden hadde noen finker funnet veien til Galapagos fra fastlandet, og siden da hadde tilfeldige mutasjoner dukket opp i fuglegene og disse endringen ble objekt for naturlig seleksjon. Som følge av dette scenariet skulle de 14 artene ha oppstått som da fantes på øyene. Splittelsen i flere arter antas å ha tatt ca. 2.3 mill. år (Sato et al, 2001). Det har ingen historiske data om når fuglene kom til øyene, så dagens konvensjonelle visdom om saken er ikke mer enn spekulasjoner. I 1967 ble omtrent 100 identiske finker flyttet fra U.S. Government Bird Reservation i midten av Stillehavet, og ble tatt med ca 500 km. avgårde til fire små atoll-øyene, som lå ca. 15 km. fra hverandre. Fuglene ble sluppet på én av øyene, og hadde snart spredt seg til alle fire. 17 år senere da fuglene ble sjekket, ble det funnet at de hadde en variasjon av nebbformer, og var tilpasset til ulike nisjer både i oppførsel og tilhørende muskler. Dette var en turbo-versjon av det konvensjonelle Galapagos-scenariet. På mindre enn 17 år, hadde finkene tilpasset seg ulike nisjer. Om denne oppdelingen ulike arter hadde funnet sted på under 20 år, hvorfor skulle det ta mer enn 2 millioner år i Darwins tilfelle.
Som kjente besøkte Ch. Darwin Galapagos under sin tur med H.M.S. Beagle. På øyene fant han finker som ikke så ut som finkene han hadde sett tidligere. Ekspertstudier av kolleger i London bekreftet dem å være nye finke-arter. Darwin teoretiserte at for lenge siden hadde noen finker funnet veien til Galapagos fra fastlandet, og siden da hadde tilfeldige mutasjoner dukket opp i fuglegene og disse endringen ble objekt for naturlig seleksjon. Som følge av dette scenariet skulle de 14 artene ha oppstått som da fantes på øyene. Splittelsen i flere arter antas å ha tatt ca. 2.3 mill. år (Sato et al, 2001). Det har ingen historiske data om når fuglene kom til øyene, så dagens konvensjonelle visdom om saken er ikke mer enn spekulasjoner. I 1967 ble omtrent 100 identiske finker flyttet fra U.S. Government Bird Reservation i midten av Stillehavet, og ble tatt med ca 500 km. avgårde til fire små atoll-øyene, som lå ca. 15 km. fra hverandre. Fuglene ble sluppet på én av øyene, og hadde snart spredt seg til alle fire. 17 år senere da fuglene ble sjekket, ble det funnet at de hadde en variasjon av nebbformer, og var tilpasset til ulike nisjer både i oppførsel og tilhørende muskler. Dette var en turbo-versjon av det konvensjonelle Galapagos-scenariet. På mindre enn 17 år, hadde finkene tilpasset seg ulike nisjer. Om denne oppdelingen ulike arter hadde funnet sted på under 20 år, hvorfor skulle det ta mer enn 2 millioner år i Darwins tilfelle.
Bilde 5. Darwins største bevis på evolusjon -hvor dypt fundert var det?
Det biokjemiske signalet som gir nebbet ulik form og fasong, er funnet å være proteinet Bmp4 (bone morphogenic protein 4). Dess mer Bmp4 som dannes i embryo-fasen, desto bredere og dypere blir fuglens nebb (Abzhanov et al, 2004). Om forfatterens antydninger er korrekte, så virker hormonene som trigges av miljømessige endringer vekstfaktorer til å forme finkens nebb. Evolusjon av finkens nebb ga ingen indikasjoner på oppbygning av informasjon {-og i Darwins tilfelle vet vi at senere da klimaet endret seg, gikk nebbstørrelsen tilbake til det normale igjen-oversetters tilføyelse}.
Karakter omflytting (character displacement) er et artsdannelses-fenomen i hvilket to arter som synes å være like når de lever atskilt, vil skille lag og bli ulike når de deler samme territorium (Brown og Wilson, 1956). Denne evnen synes designet for å hjelpe å redusere konkurransen mellom artene. Forskjellene som utviklet seg mellom dem er formentlig genetisk basert. Et slående eks. på karakter omflytting der forskjellen mellom to arter utviklet seg i løpet av ett år, etter at de to hadde levd sammen i 22 år (Grant og Grant, 2006). Den mellomstore bakkefinken (Geospiza fortis) var eneste fink på en uforstyrret Galapagos øy, inntil 1982. Da ankom tre hanner og hunner av den store bakkefinken (Geospiza magnirostris) ankom. Etter denne ankomsten ble nebbene til G. fortis målt. Gjennomsnittlig nebbstørrelse holdt seg konstant inntil 2004, da det plutselig foregikk en betydelig forminking i nebbstørrelse i løpet av ett år. (Reduksjonen var 4*ensidig 95% konfidens-grense, på estimatet for gjennomsnittsverdien i 1973.) Endringen var tilpasset i og med at den tillot G. fortis å spise på små såkorn, for å unngå konkurranse med den større og nå vel-etablerte G. magnirostris. Omgivelsesendringen var konkurranse fra eksisterende artene. Stimulus fra konkurransen bygde seg gradvis opp fra 1982. I løpet av de 7 siste årene bygde den nye bestanden (G.magnirostris) seg opp til å bli så godt som jamstor med den opprinnelige (G. fortis). Det plutselige evolusjonære fenomet som fulgte i 2004, peker sikkert mot en ikke-tilfeldig genetisk endring, trigget av omgivelsesendringer.
 Andre eks. på karakter omflyttings evolusjon som skjer raskt, er kroppstørrelse på enkelte og par av firfisler på små øyer (Schoener, 1970, Losos, 1990), samt kroppstørrelse på gjørmesnegler, og underkjeve lengde på tigerbiller (Pearson, 1980). Disse eks. på rask evolusjon gir ingen indikasjon på hvordan genetisk informasjon kan bli økt. Derfor støtter de heller ikke felles avstamning. Det er mange andre eks. på rask evolusjon i litteraturen utover det som er vist her. De inkluderer utvikling av moskito-fisk i Hawai, alder, modenhetslengde, vekstrater og størrelse til avkommet (Stearns, 1983).
Andre eks. på karakter omflyttings evolusjon som skjer raskt, er kroppstørrelse på enkelte og par av firfisler på små øyer (Schoener, 1970, Losos, 1990), samt kroppstørrelse på gjørmesnegler, og underkjeve lengde på tigerbiller (Pearson, 1980). Disse eks. på rask evolusjon gir ingen indikasjon på hvordan genetisk informasjon kan bli økt. Derfor støtter de heller ikke felles avstamning. Det er mange andre eks. på rask evolusjon i litteraturen utover det som er vist her. De inkluderer utvikling av moskito-fisk i Hawai, alder, modenhetslengde, vekstrater og størrelse til avkommet (Stearns, 1983).
Utvikling av 'soapberry bug', se Bilde 6. Jadera-haematoloma
-dens nese-lengde og utvikling (Carrol et al., 1997), samt formen på gressplanter (Snaydon og Davies, 1972). Thompson gir en liste på mer enn 20 eks. på rask evolusjon som observeres i dag. Ingen av dem heller gir noen pekepinn om hvordan informasjon bygges opp, og kan dermed heller ikke tas til inntekt for for felles avstamning. Eneste fornuftige alternativer for hvordan de virker en enten at de allerede var i populasjonen eller at de ble tydelige gjennom en omgivelses-endring, som beskrevet i en Ikke-tilfeldig evolusjons-mekanisme, hvorav ingen gir støtte for felles avstamning.
Mikroevolusjon og Makroevolusjon
 Termene mikroevolusjon og makroevolusjon benyttes for å skjelne små evolusjonære endringer fra større. Mikroevolusjonære endringer er arvelige endringer som kan bli observert i laboratoriet eller i naturen, og endringene er som sagt vanligvis små. Makroevolusjonære endringer er ikke direkte observerbare, men blir avledet ut fra fossiler eller molekylære data. Det sies at makroevolusjon tar lange tidsperioder og leder til nye kategorier av dyr, vanligvis over arts-nivået. Denne type av evolusjon kan ikke observeres, men bare deduseres da påstanden er at nye ordener eller klasser av dyr dannes over lange tidsperioder. I begynnelsen av det 20.århundre var det noe uenighet m.h.t. hvorvidt makroevolusjon var noe kvalitativt forskjellig fra mikroevolusjon (Hull, 1970). Noen følte at store evolusjonære endringer var resultat av plutselige endringer i kroppsform og ikke hadde noe å gjøre med naturlig seleksjon i neo-darwinistisk teori. Den foreslåtte prosessen som skulle produsere disse store evolusjonære endringene, uavhengig av naturlig seleksjon, ble kalt ortogenese. Men ved slutten av det 20.århundre, gikk konsensus hos evolusjonære biologer helt mot ortogonese (gresk ord for 'rettlinjet' og 'opprinnelse').
Termene mikroevolusjon og makroevolusjon benyttes for å skjelne små evolusjonære endringer fra større. Mikroevolusjonære endringer er arvelige endringer som kan bli observert i laboratoriet eller i naturen, og endringene er som sagt vanligvis små. Makroevolusjonære endringer er ikke direkte observerbare, men blir avledet ut fra fossiler eller molekylære data. Det sies at makroevolusjon tar lange tidsperioder og leder til nye kategorier av dyr, vanligvis over arts-nivået. Denne type av evolusjon kan ikke observeres, men bare deduseres da påstanden er at nye ordener eller klasser av dyr dannes over lange tidsperioder. I begynnelsen av det 20.århundre var det noe uenighet m.h.t. hvorvidt makroevolusjon var noe kvalitativt forskjellig fra mikroevolusjon (Hull, 1970). Noen følte at store evolusjonære endringer var resultat av plutselige endringer i kroppsform og ikke hadde noe å gjøre med naturlig seleksjon i neo-darwinistisk teori. Den foreslåtte prosessen som skulle produsere disse store evolusjonære endringene, uavhengig av naturlig seleksjon, ble kalt ortogenese. Men ved slutten av det 20.århundre, gikk konsensus hos evolusjonære biologer helt mot ortogonese (gresk ord for 'rettlinjet' og 'opprinnelse').
Bilde 7. Illustrasjon av makro-evolusjon
Termen makroevolusjon ble introdusert i 1927 av den russiske genetikeren Y. Filipchenko (1882-1930) i boka 'Variabilitet og variasjon' (Erwin, 2004). Han opprettholdt at mikroevolusjon kunne produsere evolusjon opp til dannelse av nye arter, men evolusjon over det nivået måtte involvere nye prosesser. Hans elev Th. Dobzhansky (1900-1975), en verdenskjent genetiker, var uenig med sin mentor og mente at makroevolusjon ikke var annet enn akkumulering av en lang suksesjon av mikroevolusjonære begivenheter. E.R. Dunn (1894-1956) en ledende herpetologo, var enig med Dobzhansky. Han skrev: Ingen bevis for en skjelning mellom mikroevolusjon og makroevolusjon finnes til stede. (Dunn, 1943). E.C.Olson (1910-1993), en ledende virveldyr-paleontolog, skrev at rådende oppfatning var at makroevolusjon ikke var annet enn en lang sekvens av mikroevolusjonære endringer, og at ingen nye prinsipper var involvert. Men mikroevolusjon og makroevolusjon var produkter av tilfeldige mutasjoner og naturlig seleksjon (Olson, 1965). Det er også nåværende konsensus mellom neo-darwinister at makroevolusjon bare er en utvidelse av mikroevolusjonære endringer. Argumenter som er benyttet her, tilbyr en annen type løsning. Emnet er omstridt fordi det ikke tilbys noe bevis for makroevolusjon. Det finnes bare bevis for mikroevolusjon.
Darwins utviklingslære gir en forklaring på hvordan alle levende vesener kan være i slekt med hverandre. Fra et felles opphav har et mangfold av arter blitt til gjennom gradvise, kontinuerlige endringer over milliarder av år. Men fossilene utfordrer Darwin. Ifølge dem er det vanlig at arter nesten ikke endrer seg over millioner av år. Dette er hva evolusjonsbiologene kaller stasis. Hva skjedde med evolusjonen?
Store, raske endringer
Den berømte paleontologen Stephen Jay Gould kom sammen med sin kollega Niels Eldredge med en forklaring som han kalte punktuert likevekt. Forklaringen går ut på at arter ikke endrer seg nevneverdig etter at de har oppstått i fossilrekken, og at større evolusjonære endringer foregår i løpet av relativt korte tidsperioder. Disse endringene forekommer som regel i sammenheng med dannelse av nye arter ved tilfeller av geografisk isolasjon. Det kan for eksempel være en gruppe organismer som blir adskilt etter et vulkanutbrudd og begynner å utvikle seg uavhengig av hverandre. Hvor ble det av de gradvise endringene? Må vi revurdere Darwins beskrivelse av evolusjon?
Bilde 8. Kjetil Lysne Voje -om stasis-problematikk
 Paradokset
Paradokset
I dag observerer forskere at evolusjon foregår med et visst tempo. - Hos nålevende arter ser vi stort potensial for evolusjon i form av genetisk variasjon. I tillegg ser vi gradvise endringer i naturlige populasjoner, sier Kjetil Lysne Voje, evolusjonsbiolog ved Universitetet i Oslo. Vi sitter igjen med et paradoks: Stasis-paradokset, som har sysselsatt mange evolusjonsbiologer i å forsøke å finne en forklaring. Voje er en av disse. I et foredrag på Realfagsbiblioteket på Universitetet i Oslo 27.8.2015, presenterte han stasis-paradokset og fortalte at det allerede er utarbeidet flere forslag til løsninger på mysteriet. Den mest utbredte forklaringen er stabiliserende seleksjon. Det vil si at en art som allerede er ganske godt tilpasset sitt miljø vil ha en fordel av ikke å endre seg så mye. Endringer kan rett og slett være en ulempe, og stabiliserende seleksjon vil operere vekk "avvik" fra den velfungerende formen. - Stabiliserende seleksjon er en veldig god forklaring på stasis, siden den sørger for at en art ikke endrer seg. Men forklaringen har problemer, for det er vanskelig å tenke seg at hva som er en optimal form ikke skal ha endret seg på mange millioner år, forteller Voje. En organismes optimale form er til enhver tid avhengig av omgivelsene. Et miljø i stadig endring, og konkurranse med andre arter gjør det lite sannsynlig at hva som er "optimalt" for en art forblir det samme over så lang tid. {Det må legges til at selv om Voje ser paradokset, ser han ikke at dette kan ha klare konsekvenser for neo-darwinismen. -oversetters kommentar.}
 Flytting og robuste arter
Flytting og robuste arter
En supplerende forklaring er nisjesporing. Det betyr rett og slett at artene søker seg til de områdene med de omstendighetene de allerede er tilpasset. Det er lettere å flytte på seg enn å evolvere. Det er også mulig at stasis skyldes det at noen arter er robuste og tåler mange ulike typer miljøer. - Krokodillen er et mulig eksempel på en slik art. Den har nesten ikke endret seg på mange millioner år. Den har kanskje ikke hatt behov for det? En annen mulighet er at enkelte arter har tilpasset seg svært stabile områder, mens andre har tilpasset seg ustabile områder. I dette tilfellet er det sannsynlig at de artene som har tilpasset seg de stabile områdene ikke har måttet endre seg så mye og eksistert lengre. Dermed er disse artene lettere å finne i fossilene.
Bilde 9. Stasis-problematikk: Krokodillen har nesten ikke utviklet seg på millioner av år
Begrensende gener
Beskrankninger er et fenomen som kan spille en stor rolle i det å forklare stasis. En beskrankning er noe som begrenser hvordan en organisme påvirkes av seleksjon. Det vil si at det kan hende en ønskelig form blir en umulighet på grunn av genetiske begrensninger. Ett gen påvirker gjerne flere egenskaper samtidig. For eksempel er armlengde sterkt genetisk koblet til beinlengde. Om man da ser for seg et miljø der lengre armer var en fordel, men lange ben var en stor ulempe, så ville utviklingen av lange armer forhindres. Genetiske beskrankninger avgjør altså hvor mye en art kan endre seg i forhold til hvor mye den burde endre seg.
- Sannsynligvis er det ikke bare én årsak til stasis. Flere forklaringer må nok tas i betraktning for å løse paradokset. Voje jobber selv med flere mulige forklaringer..
(Referanse: Kjetil Lysne Voje. Stasis-paradokset i evolusjon. Foredrag, Universitetet i Oslo, 27.8.2015)
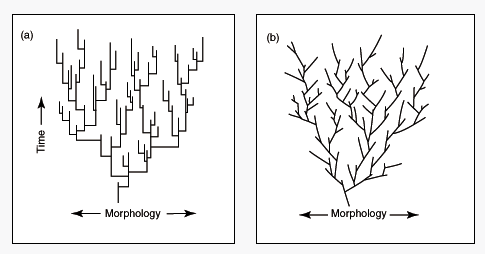 Punktert likevekt (Punctuated Equilibrium)
Punktert likevekt (Punctuated Equilibrium)
I 1972, foreslo paleontolog N. Eldridge og S.J. Gould (1941- 2002) en teori som de kalte Punktert likevekt (Punctuated Equilibrium) for å gjøre rede for makroevolusjonære endringer fortolket fra fossilregisteret. (Eldredge og Gould, 1972). De gikk mot det da fremherskende syn at mangel på mellomformer skyldes manglende fossilmateriale. Hovedtyngden av deres teori gikk på at mangelen på fossildata for gradvise evolusjonære endringer, er reell. De holdt fast på at fossilregisteret viste lange perioder med liten eller ingen endring (stasis), punktert av korte utbrudd av rask evolusjon. Disse paleontologene argumenterte for at disse eks. på stasis og likevekt i fosssilregisteret måtte begrunnes og gjøres rede for, og ikke bli oppgis som mangler i fossilregisteret, slik det hadde blitt gjort for vane siden Darwins tid. Alle darwinister er enige om at store populasjoner ikke så lett gjennomgår evolusjonære endringer, spesielt når omgivelsene er stabile. Denne stabiliteten kan forklare de mange eksemplene på stasis i fossilregisteret.
Eldredge og Gould forklarer eks. på plutselige forekomster av nye arter som resultat av separat, ikke-overlappende artsdannelse (allopatric speciation). Den tenker de skal foregå slik: En liten andel av en populasjon i dens periferi, atskiller seg fra hovedpopulasjonen.Den vesle populasjonen vil sannsynligvis ha et ikke-representativt utvalg av genetisk sammensetning fra foreldre-generasjonen. Det forårsaker at den lille populasjonen drifter avgårde fra hovedpopulasjonen. Om det varer i tilstrekkelig lang tid, kan det føre til dannelse av en ny art. Om den skulle vende tilbake til opprinnelsesstedet, ville den ikke lenger være i stand til å pare seg med den opprinnelige varianten, grunnet reproduktiv isolasjon. F.eks. via at kromosomene hadde utviklet seg til et punkt hvor hybridene ikke ville være i stand til å fungere skikkelig, eller at den hadde utviklet nye  preferanser m.h.t. parring. Om da den nye arten skulle ha en fordel overfor den gamle, ville den nye arten erstatte den gamle via naturlig seleksjon, og fossilregisteret ville vise en lang periode med likevekt (stasis) avløst av en plutselig forandring, i forhold til geologisk tid, til den nye.
preferanser m.h.t. parring. Om da den nye arten skulle ha en fordel overfor den gamle, ville den nye arten erstatte den gamle via naturlig seleksjon, og fossilregisteret ville vise en lang periode med likevekt (stasis) avløst av en plutselig forandring, i forhold til geologisk tid, til den nye.
Bilde 10. Punktert likevekt
Teorien om avbrutt likevekt endrer således fokus fra konkurranse mellom individer, til konkurranse mellom arter. De vektlegger seleksjon mellom arter som et nytt trekk, som de innrømmer går noe utenom neo-darwinsk teori. Eldredge og Gould har gjort et viktig bidrag i å påpeke at fossilregisteret viser lange perioder med likevekt, punktert av korte perioder med punktering av denne. Deres hypotese om separat, ikke-overlappende artsdannelse (allopatric speciation), synes imidlertid ikke helt å være tilfredsstillende. Dr. Spetners forslag går på at korte frambrudd av artsdannelse, mer sannsynlig stammer fra den type artsdannelse som er eksemplifisert ovenfor (sympatric speciation). Noen av dem representerer jo forekomster av omgivelses-baserte genetiske og fenotype-endringer, som er blitt observert å inntreffe i endog samme generasjon. Dette forslaget er enklere ( Ockhams barberkniv) i og med at det ikke krever noen ut-og tilbake-vandring. Det støtter seg til den veletablerte mekanismen til omgivelses-stimuli som trigger tilpassede genetiske endringer (se kap. 2). Da kan slike endringer inntreffe raskt, også i forhold til menneskelige tidsbegreper. Da Eldredge og Gould foreslo sin hypotese var rådende oppfatning at nye artsdannelser skjedde ved separat, ikke-overlappende, artsdannelse (allopatric speciation). E. Mayr, som var nestor i det 20.århundres biologi, endret oppfatning grunnet nye bevis for samtidig (sympatric) artsdannelse. (Hölldobler, 2004). Ut fra det vi vet nå, og fra diskusjonen om en Ikke-tilfeldig evolusjonsmekanisme, så er er samtidig (sympatric) artsdannelse, ut fra en Ikke-tilfeldig evolusjonsmekanisme en mer sannsynlig forklaring på punktering av likevekt-fenomenet. Tilhengere av felles avstamning har ikke i tilstrekkelig grad adressert problemet med hvordan informasjon i komplekse, nålevende organismer ble bygd opp fra antatte fjerne stamfedre. Uten en slik forklaring, er det heller ingen gyldig teori om felles avstamning.
Det forventes som vanlig ingen positive reaksjoner fra evolusjonisthold, selv om en her kommer med konkrete alternativer. Berit Ås' berømte fem hersketeknikker -her, kan strekkes ut til et mer aggregert og strukturelt nivå hvor samfunnet som helhet benytter hersketeknikker for å holde samfunnsgrupper, under glasstaket. Et glasstak flere studier bekrefter at eksisterer.
Stoffutvalg og bilder ved Asbjørn E. Lund