"Søppel DNA" lider et nederlag i det Nature artikkel finner "global funksjon" for introner i spirende gjær
 Oversatt herfra i Evolution news; 21. jan. 2019
Oversatt herfra i Evolution news; 21. jan. 2019
Bilde 1: Saccharomyces cerevisiae, by Rainis Venta [CC BY-SA 3.0], from Wikimedia Commons.
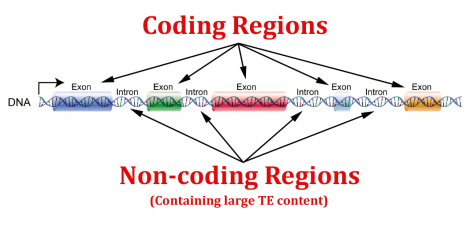
Introner er segmenter av gener som finnes i eukaryoter, som ikke kodes for proteiner. Introner fjernes fra pre-mRNA transkriptet av et proteinkompleks kalt et spliceosom, og de proteinkodende regioner (kalt eksoner) limes sammen av et annet enzym, RNA ligase.
Introner ble først identifisert i 1977 av laboratoriet av Richard Roberts og Phil Sharp (Chow et al ., 1977; Berk and Sharp, 1977; Berget and Sharp, 1977). Intronene til høyere eukaryoter kan ofte være veldig lange - i mange tilfeller spenner de over hundre tusen eller til og med millioner av baser. Lavere eukaryoter (for eksempel gjær) har en tendens til å ha kortere og færre introner. Mens de en gang var tenkt å være søppelgeneriske rester, tilbake fra millioner av evolusjonsår, har nyere bevis avslørt mange viktige  funksjoner for disse ikke-kodende regioner av gener.
funksjoner for disse ikke-kodende regioner av gener.
Det har for eksempel blitt fastslått at introner inneholder koder som er involvert i reguleringen av alternativ spleising (f.eks. Kabat et al ., 2006). Det har også vist seg at lengden på introner (og dermed tiden for å transkribere dem) kan bidra til timing-mekanismer under utvikling (Swinburne og Silver, 2010). Introner kan også kode RNA-molekyler som mikroRNA (som kreves for uttrykk av mRNA under utvikling) og små nukleolære RNA (som spiller en viktig rolle i behandlingen av ribosomale RNAer) (f.eks. Qi et al ., 2010; Kiss and Filipowicz, 1995). Mange andre funksjoner av introniske sekvenser kan diskuteres.
Introner i spirende gjær
Saccharomyces cerevisiae innehar et kompakt genom, som bare inneholder 295 introner som finnes blant 280 gener (Hooks et al ., 2014; Neuvéglise et al ., 2011). De fleste av disse intronene er kortere enn 500 nukleotider i lengde, og bare ni gjærgener har mer enn ett intron (Neuvéglise et al ., 2011; Spingola et al ., 1999).
 Det siste utgaven av Nature inneholder en artikkel om funksjonene til introner i spirende gjær (Parenteau et al ., 2019). Her er abstraktet:
Det siste utgaven av Nature inneholder en artikkel om funksjonene til introner i spirende gjær (Parenteau et al ., 2019). Her er abstraktet:
Introner er allestedsnærværende trekk ved alle eukaryotiske celler. Introner må fjernes fra fremvoksende messenger RNA gjennom spleisings-prosessen for å produsere funksjonelle proteiner. Her viser vi at den fysiske tilstedeværelsen av introner i genomet fremmer celleoverlevelse under sultforhold. Et systematisk slettingssett med alle kjente introner i spirende gjærgener indikerer at i de fleste tilfeller er celler med intron-sletting svekket, når næringsstoffer er utarmet. Denne effekten av introner under vekst er ikke knyttet til uttrykket av vertsgenet og ble reprodusert selv når oversettelse av verten mRNA ble blokkert. Transkriptomiske og genetiske analyser indikerer at introner fremmer motstand mot sult ved å øke undertrykkelsen av ribosomale protein-gener som er nedstrøms for nærings-sensende TORC1- og PKA-veier. Våre resultater avslører funksjoner av introner som kan bidra til å forklare deres evolusjonære bevaring i gener, og avdekke regulatoriske mekanismer for celle-tilpasninger til sult.
Artikkelen forklarer videre:
I denne studien presenterer vi hva som er - til vår kunnskap - den første komplette samlingen av gjærstammer hver med en sletting av en bestemt intron. Analysene av denne samlingen gir direkte bevis for globale funksjoner av introner som kan forklare deres bevaring under evolusjonen, uavhengig av funksjonen eller uttrykket for vertsgenet til intronet.
 Et bibliotek av gjærstammer
Et bibliotek av gjærstammer
Forskerne, ledet av Sherif Abou Elela fra University of Sherbrooke, konstruerte systematisk et bibliotek av gjærstammer, og slettet et ulikt intron fra hver av de 295 stammene. Resultatet av å slette introner var en reduksjon av celleveksten i nærings-fattige miljøer, selv om det ikke var mye innvirkning på celler hvis miljø ikke var utarmet av næringsstoffer. Elela og hans team fastslått at omtrent 90 prosent av introner over genomet av Saccharomyces hadde dette resultatet når de ble fjernet. I deres egne ord, "Vi konkluderer med at introner spesifikt er krevet for vekst i kulturenes stasjonære fase når næringsstoffer er utarmet."
I diskusjonen bemerker de:
Vi har vist at introner påvirker celleveksten som respons på nærings-uttørking, uavhengig av verts-genfunksjonen. Ved å slette en enkelt intron reduseres kapasiteten til celler for å motstå nærings-uttørring eller sult (figur 1, 2). Spesielt kan introner selvstendig redde defekter forårsaket av intron-sletting, selv når vertsproteinet ikke ble produsert (figur 3). Transkriptom analyser indikerer at introner fremmer motstand mot nærings utarming ved å hemme et felles sett med gener som er forbundet med oversettelse og respirasjon (figur 5). Disse intron-effektene ser ut til å koble TOR- og PKA-banene til undertrykkelsen av ribosom-biogenese, på grunnlag av næringsinnholds konsentrasjonen (figur 6 og utvidede data figur 10b, c). Sammen presenterer dataene et paradigme for intron-funksjon der tilstedeværelsen av introner direkte bidrar til cellevekst på en måte som er uavhengig av funksjonen av vertsgenet.
I samme utgave av Nature rapporterte et annet team, ledet av RNA biolog David Bartel fra Massachusetts Institute of Technology, 34 introner i Saccharomyces cerevisiae som "akkumuleres som lineære RNAer under enten mettet vekstbetingelser eller andre  påkjenninger som forårsaker langvarig hindring av TORC1, som er en sentral integrator for vekstsignalering "(Morgan et al ., 2019). Vanligvis har celler blitt analysert under logget fasevekst hvor celle-proliferasjon ikke er begrenset og deling skjer i en konstant hastighet. Siden dette vanligvis ikke reflekterer situasjonen til spirende gjær i et naturlig miljø, søkte forskerne å undersøke dens genregulering i en annen sammenheng enn den loggede fase-veksten.
påkjenninger som forårsaker langvarig hindring av TORC1, som er en sentral integrator for vekstsignalering "(Morgan et al ., 2019). Vanligvis har celler blitt analysert under logget fasevekst hvor celle-proliferasjon ikke er begrenset og deling skjer i en konstant hastighet. Siden dette vanligvis ikke reflekterer situasjonen til spirende gjær i et naturlig miljø, søkte forskerne å undersøke dens genregulering i en annen sammenheng enn den loggede fase-veksten.
Forskerne slettet en delmengde av disse intronene fra Saccharomyces- genomet ved hjelp av CRISPR, og sammenlignet de resulterende celler med villtype-celler for å bestemme effekten på cellevekst. I tråd med resultatene fra Elela og hans kollegaer oppdaget de at cellene av villtype fungerte bra i nærings-fattige miljøer, men ikke så bra da det var en overflod av ressurser. De forandrede cellene (med introner slettet) gjorde det derimot godt når det var rikelig med ressurser, men ikke når ressursene var knappe.
Overlevelse i ressurskrevne miljøer
Hvordan fremmer introner celleoverlevelse i forhold til utarmede ressurser? En mulighet, foreslått av forskerne, er at spliceosomer er isolert av stabile introner. Det vil si at spliceosom-apparatet blir 'rotet opp' av introner slik at det hindres fra å spleise nylig transkriberte introner. Forskerne foreslo en modell hvor TORC1-banen, en signalkaskade som er ansvarlig for å regulere gjærcelleveksten som respons på næringsstoff-tilgjengelighet, fører til oppbygging av introner i ressurs-utarmede miljøer. Dette ville være en fordel, siden det ville hindre at energi blir spilt på forsøk på å vokse i miljøer hvor ressursene er knappe. Faktisk er TORC1 allerede kjent for å regulere, på næringsavhengig måte, uttrykket av ribosomale proteiner (Li et al ., 2006).
 Elelas lag har utført ytterligere eksperimenter for å foreslå at i næringsdepleterte celler undertrykkes uttrykket av ribosomale proteiner som trengs for protein-oversettelse av introner. Dette antyder at intronene hemmer spleising og oversettelse, og reduserer dermed frekvensen av cellemetabolisme og energiforbruk.
Elelas lag har utført ytterligere eksperimenter for å foreslå at i næringsdepleterte celler undertrykkes uttrykket av ribosomale proteiner som trengs for protein-oversettelse av introner. Dette antyder at intronene hemmer spleising og oversettelse, og reduserer dermed frekvensen av cellemetabolisme og energiforbruk.
Alt dette bringer til minne en kommentar fra biologen John Mattick, en kritiker av søppel- DNA paradigmet. Tilbake i 2003 i Scientific American (Gibbs, 2003) skrev han:
Mangelen på å gjenkjenne de fullstendige konsekvensene av dette - særlig muligheten for at de intervenerende ikke-kodende sekvensene kan overføre parallell informasjon i form av RNA-molekyler - kan godt nedkomme som en av de største feilene i molekylærbiologiens historie.
Nettopp det.
Litteratur sitert:
Berget, SM, Sharp, PA (1977) En spleiset sekvens ved 5'-terminalen av adenovirus sent mRNA. Brookhaven Symp Biol , 29: 332-44.
Berk, AJ, Sharp, PA (1977) Dimensjonering og kartlegging av tidlige adenovirus mRNAer ved gelelektroforese av S1 endonuklease-fordøyede hybrider. Cell 12 (3): 721-32.
Chow, LT, Roberts, JM, Lewis, JB, Broker, TR (1977) Et kart over cytoplasmatiske RNA-transkripsjoner fra lytisk adenovirus type 2, bestemt ved elektronmikroskopi av RNA: DNA-hybrider. Cell , 11 (4): 819-36.
Gibbs, WW (2003) Det usete genomet: edelstener blant søppel. Vitenskapelig amerikansk 289 (5): 26-33.
Hooks, KB, Delneri, D. & Grifths-Jones, S. (2014) Intron evolution in Saccharomycetaceae. Genombiol. Evol . 6, 2543-2556.
Kabat, JL, Barberan-Soler, S., McKenna, P., Clawson, H., Farrer, T. og Zahler, AM (2006) Intronic Alternative Splicing Regulators Identifisert av Comparative Genomics in Nematodes. PLoS Computational Biology 2 (7): 734-747.
 Kiss, T. og Filipowicz, W. (1995) Genes and Development 9 (11): 1411-1424.
Kiss, T. og Filipowicz, W. (1995) Genes and Development 9 (11): 1411-1424.
Li, H., Tsang, CK, Watkins, M., Bertram, PG og Zheng, XF (2006). Næringsstoff regulerer Tor1-nukleær lokalisering og forening med rDNA-promotor. Nature 442, 1058-1061.
Morgan, JT, Fink, GR og Bartel, DP (2019) Spente lineære introner regulerer veksten i gjær. Naturen .
Neuvéglise, C., Marck, C. og Gaillardin, C. (2011). Intronomet av spirende gjær. Comptes Rendus Biologies 334: 662-670.
Parenteau, J. et al. (2019), "Introns er mediatorer av celle respons på sult," Nature.
Spingola, M., Grate, L., Haussler, D. & Ares, M. Jr. (1999). Genomfattende bioinformatisk og molekylær analyse av introner i Saccharomyces cerevisiae. RNA 5, 221-234.
Monteys, AM et al . (2010) Struktur og aktivitet av formodede introniske miRNA promotorer. RNA 25 (2): 495-505.
Swinburne, IA og Silver, PA (2008) Intronforsinkelser og transkriptionell timing under utvikling. Utviklingscell 14 (3): 324-330.
Qi, Y. et al. (2010) High-Throughput Sequencing of MicroRNAs in Adenovirus Type 3 Infected Human Larynge Epithelial Cells. Journal of Biomedicine and Biotechnology 2010.
Oversettelse og bilder ved Asbjørn E. Lund