LIVETS OPPRINNELSE
 LIVET BURDE IKKE EKSISTERE. Så mye kjenner vi til fra kjemi. I motsetning til livets allestedsnærværelse på jorden, virker livløsheten til andre planeter langt bedre fornuftig kjemiskisk sett. Syntetiske kjemikere vet hva som trengs for å bygge bare én molekylær forbindelse. Forbindelsen må utformes og bli stereokjemisk kontrollert. Utbytteoptimalisering, rensing og karakterisering er nødvendig. En omfattende forsyning er nødvendig for å kontrollere syntesen fra start til slutt. Intet av dette er lett. Få forskere fra andre fagområder forstår hvordan molekyler syntetiseres.
LIVET BURDE IKKE EKSISTERE. Så mye kjenner vi til fra kjemi. I motsetning til livets allestedsnærværelse på jorden, virker livløsheten til andre planeter langt bedre fornuftig kjemiskisk sett. Syntetiske kjemikere vet hva som trengs for å bygge bare én molekylær forbindelse. Forbindelsen må utformes og bli stereokjemisk kontrollert. Utbytteoptimalisering, rensing og karakterisering er nødvendig. En omfattende forsyning er nødvendig for å kontrollere syntesen fra start til slutt. Intet av dette er lett. Få forskere fra andre fagområder forstår hvordan molekyler syntetiseres.
Syntetiske begrensninger må tas i betraktning når man vurderer prebiotisk fremstilling av de fire klassene av forbindelser som er nødvendige for livet: aminosyrene, nukleotidene, sakkaridene og lipidene. (1) Det neste nivået utover syntese innebærer komponentene som trengs for bygging av nanosystemer, som deretter settes sammen i et mikrosystem. Sammensatt av mange nanosystemer, er cellen et naturens grunnleggende mikrosystem. Hvis de første cellene var relativt enkle, krevde de fremdeles minst 256 proteinkodende-gener. Dette kravet er så nært som absolutt som vi finner i syntetisk kjemi. En bakterie som koder for 1.354 proteiner inneholder en av de minste genomene som er kjent pr.d.d. (2)
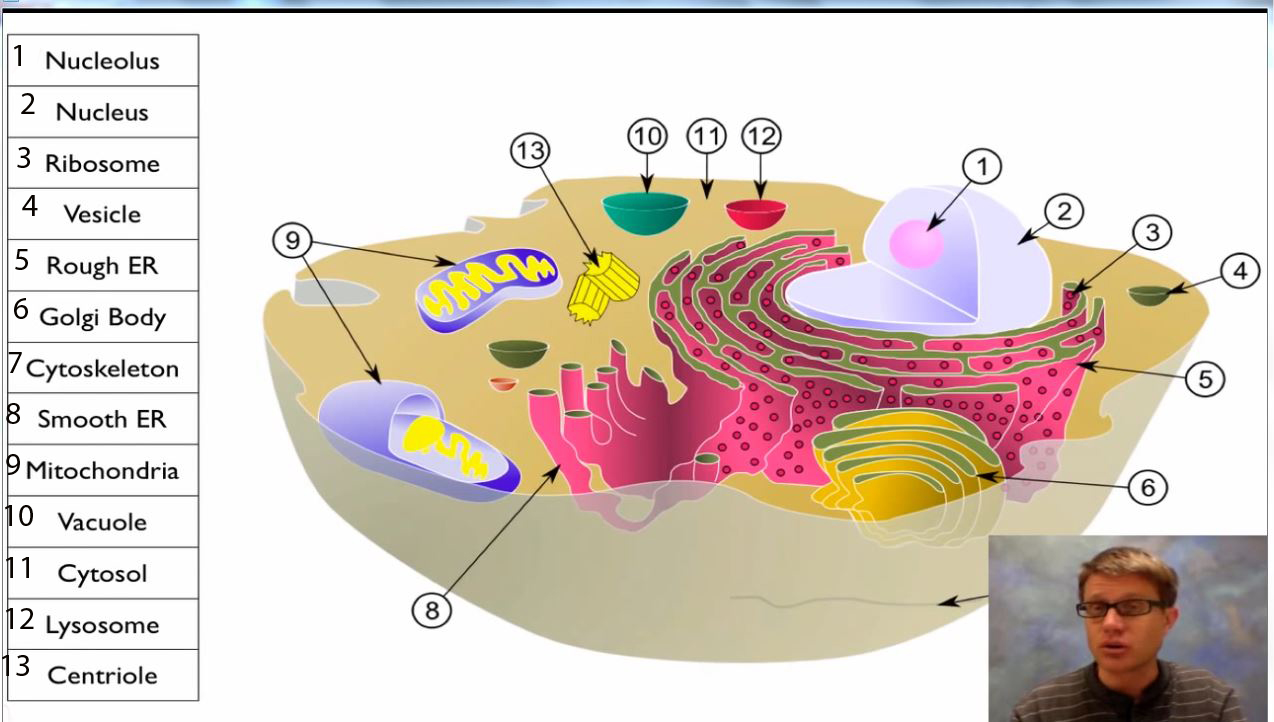 Vurder følgende tankeeksperiment. La oss anta at alle molekylene vi tror kan være nødvendige for å konstruere en celle, er tilgjengelige i de nødvendige kjemiske og stereokjemiske renheter. La oss anta at disse molekylene kan skilles og leveres til et velutstyrt laboratorium. La oss også anta at de millioner av artikler som består av den kjemiske og biokjemiske litteraturen, er lett tilgjengelige.
Vurder følgende tankeeksperiment. La oss anta at alle molekylene vi tror kan være nødvendige for å konstruere en celle, er tilgjengelige i de nødvendige kjemiske og stereokjemiske renheter. La oss anta at disse molekylene kan skilles og leveres til et velutstyrt laboratorium. La oss også anta at de millioner av artikler som består av den kjemiske og biokjemiske litteraturen, er lett tilgjengelige.
Hvordan kan vi bygge en celle?
Det er ikke nok å ha kjemikaliene til stede. Forholdet mellom nukleotidene og alt annet må spesifiseres, og for dette er kodende informasjon avgjørende. DNA og RNA er de primære informasjonsbærerne i cellen. Uansett hvilket medium livet kan ha benyttet i begynnelsen, så måtte informasjonen komme fra et sted. En streng av nukleotider koder ikke i seg selv noe. La oss anta at DNA og RNA er tilgjengelige i hvilken rekkefølge vi ønsker.
En celle, som definert i syntetiske biologiske termer, er et system som kan vedlikeholde iongradienter, fange og behandle energi, lagre informasjon og mutere. (3) Kan vi bygge en celle fra råvarene? (4) Vi er tross alt syntetiske kjemikere. Hvis vi ikke kan gjøre det, kan ingen. Lipider av passende lengde kan spontant danne lipid bilag.
Molekylærbiologi lærebøker sier så mye. En lipid dobbeltlagsboble kan inneholde vann, og var en sannsynlig forløper til den moderne cellemembranen. (5) Lipid ansamling i en lipid bilag-membran kan enkelt fremprovoseres ved rystelse eller lydbehandling i et laboratorium.
Et voilà . Det nødvendige lipid-dobbeltlaget dannes deretter. Ikke sant?
Ikke så fort. Noen problemer bør gi oss en pause: (6)
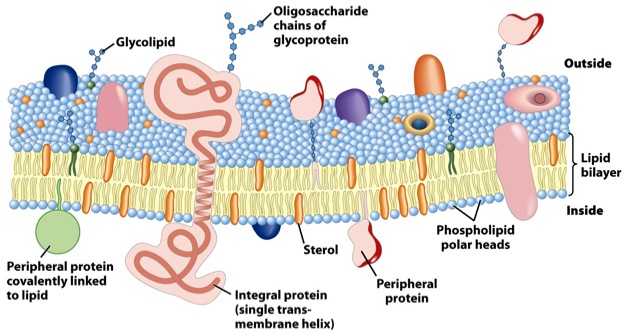 Forskere har identifisert tusenvis av forskjellige lipidstrukturer i moderne cellemembraner. Disse inkluderer glycerolipider, sfingolipider, steroler, prenoler, sakkarolipider og polyketider. (7) Av denne grunn er valg av dobbeltlags-blanding for vårt syntetiske membranmål langt fra rett fram. Når man lager syntetiske vesikler-syntetiske lipid-dobbeltlagsmembraner, kan blandinger av lipider bør noteres, destabilisere systemet.
Forskere har identifisert tusenvis av forskjellige lipidstrukturer i moderne cellemembraner. Disse inkluderer glycerolipider, sfingolipider, steroler, prenoler, sakkarolipider og polyketider. (7) Av denne grunn er valg av dobbeltlags-blanding for vårt syntetiske membranmål langt fra rett fram. Når man lager syntetiske vesikler-syntetiske lipid-dobbeltlagsmembraner, kan blandinger av lipider bør noteres, destabilisere systemet.
Lipid dobbeltsidige lag omgir subcellulære organeller, som kjerner og mitokondrier, som selv er nanosystemer og mikrosystemer. Hver av disse har sin egen lipid-sammensetning.
Lipider har en ikke-symmetrisk fordeling. Ytre og indre flater av lipid bi-lagene er kjemisk ikke-ekvivalente, og kan ikke byttes om på.
Lipidene er bare begynnelsen. Protein-lipidkomplekser er de nødvendige passive transportstedene og pumper aktivt for gjennomstrømning av ioner og molekyler gjennom dobbeltlags-membraner, ofte med høy spesifisitet. Noen tillater passasje for substrater inn i rommet, og andre deres utgang. Kompleksiteten øker ytterligere fordi alle lipid bi-lag har et stort antall polysakkarid (sukker)  vedheng, kjent som glykaner (kompliserte sukkerforbindelser), og sukrene er ingen spøk. Disse er viktige for nanosystem og mikrosystem-regulering. Den saklige kompleksiteten av disse sakkaridene er skremmende. Seks gjentatte enheter av sakkarid D-pyranose kan danne mer enn en trillion forskjellige heksasakkarider gjennom forgrening (konstitusjonell) og glykosidisk (stereokjemisk) mangfoldighet. (8) Tenk på bredden av biblioteket!
vedheng, kjent som glykaner (kompliserte sukkerforbindelser), og sukrene er ingen spøk. Disse er viktige for nanosystem og mikrosystem-regulering. Den saklige kompleksiteten av disse sakkaridene er skremmende. Seks gjentatte enheter av sakkarid D-pyranose kan danne mer enn en trillion forskjellige heksasakkarider gjennom forgrening (konstitusjonell) og glykosidisk (stereokjemisk) mangfoldighet. (8) Tenk på bredden av biblioteket!
Polysakkarider er de mest tallrike organiske molekylene på planeten. Deres betydning gjenspeiles i det faktum at de er produsert av og er avgjørende for alle naturlige systemer. Hver cellemembran er belagt med et komplekst utvalg av polysakkarider, og alle celle-til-celle-interaksjoner finner sted gjennom sakkarid-deltakelse på den dobbelte lipid-membranoverflaten. Eliminasjon av enhver klasse av sakkarider fra en organisme resulterer i dens død, og hver cellulær dysfunksjon involverer sakkarider.
I en rapport med tittelen "Transforming Glycoscience", konstaterte USAs nasjonale forskningsråd nylig at, svært lite er kjent om glykaners diversifisering under utviklingen. Over tre milliarder år med evolusjon har unnlatt å generere noen form for levende celle som ikke er dekket av et tett og komplekst utvalg av glykaner. (9)
I tillegg observerte Vlatka Zoldos, Tomislav Horvat og Gordan Lauc: "En egenart ved glykan-deler av glykoproteiner er at de ikke 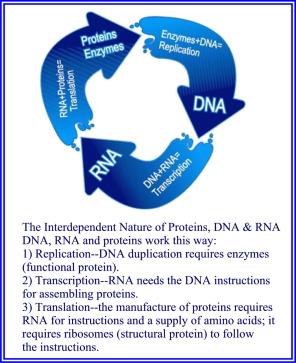 syntetiseres ved hjelp av en direkte genetisk mal. I stedet oppstår de fra aktiviteten til flere hundrevis av enzymer organisert i komplekse veier." -her (10)
syntetiseres ved hjelp av en direkte genetisk mal. I stedet oppstår de fra aktiviteten til flere hundrevis av enzymer organisert i komplekse veier." -her (10)
Sakkarider er informasjonsrike molekyler. Glykosyltransferaser koder informasjon til glykaner og sakkaridbindende proteiner og dekoder informasjonen lagret i glykanstrukturer. Denne prosessen gjentas i henhold til polysakkarid forgrenings- og koblingsmønstre. (11) Sakkarider koder og overfører informasjon lenge etter sin første enzymatiske konstruksjon. (12) Polysakkarider bærer mer potensiell informasjon enn noe annet makromolekyl, inkludert DNA og RNA. Av denne grunn viser lipidassosierte polysakkarider seg gåtefulle. (13)
Cellular og organelle dobbeltsidige lag, som en gang var tenkt på som enkle vesikler, er alt annet enn det. De er svært funksjonelle portvoktere. I kraft av deres glykaner blir lipidbi-lagene enorme banker med lagret, lesbar og omskrivbar informasjon. Lydbehandling av noen få tilfeldige lipider, polysakkarider og proteiner i et laboratorium vil ikke gi cellulære lipid-dobbeltlags-membraner.
Mine venner, mine likemenn, med disse kompleksitetene i tankene, hvordan kan vi bygge mikrosystemet til en enkel celle? Ville vi kunne bygge endog lipid bi-lagene? Disse mindre cellulære mikrosystemene, som igjen består av tusenvis av nanosystemer, er utenfor vår forståelse. Likevel er vi ledet til å tro at for 3,8 milliarder år siden kunne de nødvendige forbindelsene bli funnet i en eller annen hule eller undervanns-sjakt, og på en eller annen måte samlet seg inn i den første cellen.
Kunne tiden virkelig ha bevirket slik magi?
Mange av de molekylære strukturer som trengs for livet, er ikke termodynamisk favorisert av deres syntese. Dannet ved formose-reaksjonen, gjennomgikk sakkaridene videre kondensering under de samme reaksjons-betingelser de ble dannet i. Resultatet er polymert materiale, for ikke å nevne stereo-tilfeldighet i hvert stereo-oppvekstsenter, derfor dobbelt ubrukelige(!) (14) Tiden er fienden. Reaksjonen må stoppes kort tid etter at det ønskede produktet er nådd. Hvis vi går tom for syntetiske mellomprodukter i laboratoriet, må vi gå tilbake til begynnelsen. Naturen holder ikke en lab-notatbok. Hvordan bringer hun opp mer materiale fra 'lageret'?
Hvis man forstår termodynamikkens andre lov, slik noen fysikere gjør, (15) Så "kan du begynne med en tilfeldig klump av atomer, og hvis du lyser på den lenge nok, bør det ikke være så overraskende at du får en plante." (16) Lysets vekselvirkning med små molekyler er godt forstått. Forsøket er utført. Resultatet er kjent. Uansett lysets bølgelengde, dannes ingen plante noen gang.
 Vi syntetiske kjemikere bør si det åpenbare. Oppdukkingen av livet på jorden er et mysterium. Vi er ikke i nærheten av å løse dette problemet. Forslagene som tilbys til nå for å forklare livets opprinnelse, gir ingen vitenskapelig mening.
Vi syntetiske kjemikere bør si det åpenbare. Oppdukkingen av livet på jorden er et mysterium. Vi er ikke i nærheten av å løse dette problemet. Forslagene som tilbys til nå for å forklare livets opprinnelse, gir ingen vitenskapelig mening.
Utover vår planet, er alle de andre som har blitt undersøkt livløse, et resultat i tråd med våre kjemiske forventninger. Lovene i fysikkens og kjemiens periodiske tabeller er universelle, noe som tyder på at livet basert på aminosyrer, nukleotider, sakkarider og lipider er en anomali. Livet burde ikke eksistere noe som helst sted i vårt univers. Livet skulle ikke engang eksistere på jordens overflate. (17)
Referanser:
1. See James Tour, "Animadversions of a Synthetic Chemist," Inference: International Review of Science 2, no. 2 (2016); James Tour, “Two Experiments in Abiogenesis,” Inference: International Review of Science 2, no. 3 (2016).
2. Se Wikipedia, "Minimal Genome."
3. David Dearner, "A Giant Step Towards Artificial Life?" Trends in Biotechnology 23, no. 7 (2008): 336-38, doi:10.1016/j.tibtech.2005.05.008.
4. En liten bit dette målet ble oppnådd da et syntetisk genom ble satt inn i en vertscelle hvorfra det originale genomet var fjernet. Bi-lags rmembranen til vertscellen og alle dens cytoplasmatiske bestanddeler var allerede opprettet av naturlige biologiske prosesser. Se Daniel Gibson et al., "Creation of a Bacterial Cell Controlled by a Chemically Synthesized Genome," Science 329, no. 5,987 (2010): 52-56, doi:10.1126/science.1190719.
5. Bruce Alberts et al., Molecular Biology of the Cell, 4th ed. (New York: Garland Science, 2002).
6. Se F. Xabier Contreras et al., "Molecular Recognition of a Single Sphingolipid Species by a Protein's Transmembrane Domain," Nature 481 (2012): 525-29, doi:10.1038/nature10742; Yoshiyuki Norimatsu et al., "Protein-Phospholipid Interplay Revealed with Crystals of a Calcium Pump," Nature 545 (2017): 193-98, doi:10.1038/nature22357.
7. Se Lipidomics Gateway, "LIPID MAPS Structure Database."
8. Roger Laine, "Invited Commentary: A Calculation of All Possible Oligosaccharide Isomers Both Branched and Linear Yields (1.05 ganger 1012) Structures for a Reducing Hexasaccharide: The Isomer Barrier to Development of Single-Method Saccharide Sequencing or Synthesis Systems," Glycobiology 4, no. 6 (1994): 759-67, doi:10.1093/glycob/4.6.759.
9. National Research Council, Transforming Glycoscience: A Roadmap for the Future (Washington, DC: The National Academies Press, 2012), 72, doi:10.17226/13446.
10. Vlatka Zoldos, Tomislav Horvat and Gordan Lauc, "Glycomics Meets Genomics, Epigenomics and Other High Throughput Omics for System Biology Studies," Current Opinion in Chemical Biology 17, no. 1 (2012): 33-40, doi:10.1016/j.cbpa.2012.12.007.
11. Adapted from Maureen Taylor and Kurt Drickamer, Introduction to Glycobiology-her (Oxford: Oxford University Press, 2006).
12. Gordan Lauc, Aleksandar Vojta and Vlatka Zoldos, "Epigenetic Regulation of Glycosylation Is the Quantum Mechanics of Biology," -her Biochimica et Biophysica Acta - General Subjects 1,840, no. 1 (2014): 65-70, doi:10.1016/j.bbagen.2013.08.017.
13. Claus-Wilhelm von der Lieth, Thomas Luetteke, and Martin Frank, eds., Bioinformatics for Glycobiology and Glycomics: An Introduction (Chichester: Wiley-Blackwell, 2009).
14. James Tour, "Animadversions of a Synthetic Chemist," Inference: International Review of Science 2, no. 2 (2016). -her
15. See Jeremy England, "Statistical Physics of Self-Replication," Journal of Chemical Physics 139 (2013), doi:10.1063/1.4818538 her; Paul Rosenberg, "God is on the Ropes: The Brilliant New Science That Has Creationists and the Christian Right Terrified," Salon, January 3, 2015.
16. Natalie Wolchover, " New Physics Theory of Life," Quanta, January 22, 2014. -her
17. The author wishes to thank Anthony Futerman of the Weizmann Institute and Russell Carlson of the University of Georgia for information on lipids and saccharides, respectively.
Oversettelse og bilder ved Asbjørn E. Lund